Download

1 / 44
450 likes | 858 Vues
This guide explores gene linkage and mapping, Mendel’s laws, chromosome loci, alleles, and genetic mapping methods. Learn how genetic polymorphisms and recombination frequencies help determine gene distances and order. Discover the significance of human pedigrees in genetic mapping.

E N D
4 Gene Linkage and Genetic Mapping
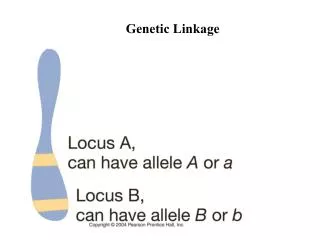
Mendel’s Laws: Chromosomes • Locus = physical location of a gene on a chromosome • Homologous pairs of chromosomes often contain alternative forms of a given gene = alleles • Different alleles of the same gene segregate at meiosis I • Alleles of different genes assort independently in gametes • Genes on the same chromosome exhibit linkage: inherited together
Gene Mapping • Gene mapping determines the order of genes and the relative distances between them in map units • 1 map unit= 1 cM (centimorgan) In double heterozyote: • Cisconfiguration = mutant alleles of both genes are on the same chromosome = ab/AB • Trans configuration = mutant alleles are on different homologues of the same chromosome = Ab/aB
Gene Mapping • Gene mapping methods use recombination frequencies between alleles in order to determine the relative distances between them • Recombination frequencies between genes are inversely proportional to their distance apart • Distance measurement: 1 map unit = 1 percent recombination (true for short distances)
Gene Mapping • Genes with recombination frequencies less than 50 percent are on the same chromosome = linked) • Linkage group = all known genes on a chromosome • Two genes that undergo independent assortment have recombination frequency of 50 percent and are located on nonhomologous chromosomes or far apart on the same chromosome = unlinked Fig. 4.7
Recombination • Recombination between linked genes occurs at the same frequency whether alleles are in cis ortransconfiguration • Recombination frequency is specific for a particular pair of genes • Recombination frequency increases with increasing distances between genes • No matter how far apart two genes may be, the maximum frequency of recombination between any two genes is 50 percent.
Gene Mapping • Recombination results from crossing-over between linked alleles. • Recombination changes the allelic arrangement on homologous chromosomes Fig. 4.4
Genetic Mapping • The map distance (cM) between two genes equals one half the average number of crossovers in that region per meiotic cell • Therecombination frequency between two genes indicates how much recombination is actually observed in a particular experiment; it is a measure of recombination • Over an interval so short that multiple crossovers are precluded (~ 10 percent recombination or less), the map distance equals the recombination frequency because all crossovers result in recombinant gametes. • Genetic map=linkage map=chromosome map
Gene Mapping: Crossing Over • Two exchanges taking place between genes, and both involving the same pair of chromatids, result in a nonrecombinant chromosomes Fig. 4.13
Gene Mapping: Crossing Over • Crossovers which occur outside the region between two genes will not alter their arrangement • The result ofdouble crossoversbetween two genes is indistinguishable from independent assortment of the genes • Crossovers involving three pairs of alleles specify gene order = linear sequence of genes
Genetic vs. Physical Distance • Map distances based on recombination frequencies are not a direct measurement of physical distance along a chromosome • Recombination “hot spots” overestimate physical length • Low rates in heterochromatin and centromeres underestimate actual physical length
Gene Mapping • Mapping function: the relation between genetic map distance and the frequency of recombination • Chromosome interference: crossovers in one region decrease the probability of a second crossover close by • Coefficient of coincidence= observed number of double recombinants divided by the expected number Interference = 1-Coefficient of coincidence
Mapping Genes in Human Pedigrees • Methods of recombinant DNA technology are used to map human chromosomes and locate genes • Genes can then be cloned to determine structure and function • Human pedigrees and DNA mapping are used to identify dominant and recessive disease genes • PolymorphicDNA sequences are used in human genetic mapping.
Genetic Polymorphisms • The presence in a population of two or more relatively common forms of a gene or a chromosome is calledpolymorphism • Changes in DNA fragment length produced by presence or absence of the cleavage sites in DNA molecules are known as restriction fragment length polymorphism (RFLP) • A prevalent type of polymorphism is a single base pair difference, simple-nucleotide polymorphism (SNP) • A genetic polymorphism resulting from a tandemly repeated short DNA sequence is called a simple sequence repeat(SSR)
RFLPs • Restriction endonucleases are used to map genes as they produce a unique set of fragments for a gene • There are more than 200 restriction endonucleases in use, and each recognizes a specific sequence of DNA bases • EcoR1cuts double-stranded DNA at the sequence 5’-GAATTC-3’ wherever it occurs
RFLPs • Differences in DNA sequence generate different recognition sequences and DNA cleavage sites for specific restriction enzymes • Two different genes will produce different fragment patterns when cut with the same restriction enzyme due to differences in DNA sequence Fig. 4.19
SNPs • Single-nucleotide polymorphisms (SNPs) are abundant in the human genome • Rare mutants of virtually every nucleotide can probably be found, but rare variants are not generally useful for family studies of heritable variation in susceptibility to disease • For this reason, in order for a difference in nucleotide sequence to be considered as an SNP, the less-frequent base must have a frequency of greater than about 5% in the human population. • By this definition, the density of SNPs in the human genome averages about one per 1300 bp
SSRs • A third type of DNA polymorphism results from differences in the number of copies of a short DNA sequence that may be repeated many times in tandem at a particular site in a chromosome • When a DNA molecule is cleaved with a restriction endonuclease that cleaves at sites flanking the tandem repeat, the size of the DNA fragment produced is determined by the number of repeats present in the molecule • There is an average of one SSR per 2 kb of human DNA
Mapping Genes in Human Pedigrees • One source of the utility of SNPs and SSRs in human genetic mapping is their high density across the genome • Additional source of utility of SSRs in genetic mapping is the large number of alleles that can be present in any human population.
Mapping Genes in Human Pedigrees • Human pedigrees can be analyzed for the inheritance pattern of different alleles of a gene based on differences in SSRs and SNPs • Restriction enzyme cleavage of polymorphic alleles that are different in RFLP pattern produces different size fragments by gel electrophoresis
Tetrad Analysis • In some species of fungi, each meiotic tetrad is contained in a sac-like structure, called an ascus • Each product of meiosis is an ascospore, and all of the ascospores formed from one meiotic cell remain together in the ascus Fig. 4.22
Tetrad Analysis • Several features of ascus-producing organisms are especially useful for genetic analysis: • They are haploid, so the genotype is expressed directly in the phenotype • They produce very large numbers of progeny • Their life cycles tend to be short
Ordered and Unordered Tetrads • Organisms like Saccharomycescerevisiae, produce unordered tetrads: the meiotic products are not arranged in any particular order in the ascus • Unordered tetrads have no relation to the geometry of meiosis. • Bread molds of the genus Neurospora have the meiotic products arranged in a definite order directly related to the planes of the meiotic divisions—ordered tetrads • The geometry of meiosis is revealed in ordered tetrads
Tetrad Analysis • In tetrads when two pairs of alleles are segregating, three patterns of segregation are possible • parental ditype (PD) = two parental genotypes • nonparental ditype (NPD) = only recombinant combinations • tetratype (TT) = all four genotypes observed Fig. 4.24
Tetrad Analysis • The existence of TT for linked genes demonstrates two important features of crossing-over • The exchange of segments between parental chromatids takes place in prophase I, after the chromosomes have duplicated • The exchange process consists of the breaking and rejoining of the two chromatids, resulting in the reciprocal exchange of equal and corresponding segments
Tetrad Analysis • When genes are unlinked, the parental ditype tetrads and the nonparental ditype tetrads are expected in equal frequencies: PD = NPD • Linkage is indicated when nonparental ditype tetrads appear with a much lower frequency than parental ditype tetrads: PD » NPD • Map distance between two genes that are sufficiently close that double and higher levels of crossing-over can be neglected, equals 1/2 x (Number TT / Total number of tetrads) x 100
Neurospora: Ordered Tetrads • Ordered asci also can be classified as PD, NPD, or TT with respect to two pairs of alleles, which makes it possible to assess the degree of linkage between the genes • The fact that the arrangement of meiotic products is ordered also makes it possible to determine the recombination frequency between any particular gene and its centromere
Tetrad Analysis: Ordered Tetrads • Homologous centromeres of parental chromosomes separate at the first meiotic division • The centromeres of sister chromatids separate at the second meiotic division • When there is no crossover between the gene and centromere, the alleles segregate in meiosis I • A crossover between the gene and the centromere delays segregation alleles until meiosis II
Tetrad Analysis: Ordered Tetrads • The map distance between the gene and its centromere equals • 1/2 x (Number of asci with second division segregation/ Total number of asci) x 100 • This formula is valid when the gene is close enough to the centromere and there are no multiple crossovers
Gene Conversion • Most asci from heterozygous Aa diploids demonstrate normal Mendelian segregation and contain ratios of 2A : 2a in four-spored asci, or 4A : 4a in eight-spored asci Occasionally, aberrant ratios are also found, such as 3A : 1a or 1A : 3a and 5A : 3a or 3A : 5a The aberrant asci are said to result from gene conversion because it appears as if one allele has “converted” the other allele into a form like itself
Gene Conversion • Gene conversion is frequently accompanied by recombination between genetic markers on either side of the conversion event, even when the flanking markers are tightly linked • Gene conversion results from a normal DNA repair process in the cell known as mismatch repair • Gene conversion suggests a molecular mechanism of recombination
Recombination • Recombination is initiated by a double-stranded break in DNA • The size of the gap is increased by nuclease digestion of the broken ends • These gaps are repaired using the unbroken homologous DNA molecule as a template • The repair process can result in crossovers that yield chiasmata between nonsister chromatids
Recombination: Holliday Model • The nicked strands unwind, switch partners, forming a short heteroduplexregion with one strand and a looped-out region of the other strand called a Dloop • The juxtaposed free ends are joined together, further unwinding and exchange of pairing partners increase the length of heteroduplex region—process of branch migration
Recombination: Holliday Model • One of two ways to resolve the resulting structure, known as a Holliday junction, leads to recombination, the other does not • The breakage and rejoining is an enzymatic function carried out by an enzyme called the Holliday junction-resolving enzyme