Download
1 / 39
410 likes | 630 Vues
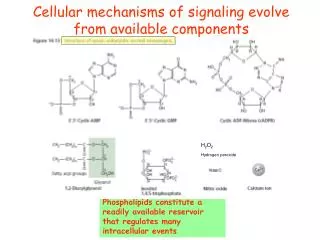
Cellular mechanisms of signaling evolve from available components. H 2 O 2 Hydrogen peroxide. Phospholipids constitute a readily available reservoir that regulates many intracellular events. PtdIns in yeast. Phospholipase C activation initiates the IP 3 pathway.

E N D
Cellular mechanisms of signaling evolve from available components H2O2 Hydrogen peroxide Phospholipids constitute a readily available reservoir that regulates many intracellular events
PhospholipaseC activation initiates the IP3 pathway Cellular mechanisms of signaling evolve from available components Phospholipids act as a readily available reservoir that regulate many intracellular events
Pathways of phosphoinositide synthesis and degradation. The phosphatidylinositol (PI) includes inositol 1-phosphate bound via its phosphate group to 1-stearoyl,2-arachidoyl diacylglycerol, prevalent in mammal cells favouring exposure of the inositol ring and its interaction with PIBMs.The 7 known PIs in eukaryotes are PI4P,PI5P,PI3P,PI45P2, PI35P2, PI34P2 and PI345P3. Each PI is indicated according to colour codes. Blue and arrows indicate routes of PI phosphorylation and dephosphorylation, respectively. Unlike phosphoinositides, the soluble inositol phosphates (IPs) can be phosphorylated in all of the six positions,giving rise to more than 60 soluble species.This is because other IP-specific enzymes are present in the cell as well as the kinases/phosphatases acting on the phosphoinositides and IPs.The PIs can be hydrolysed by PLC to generate inositol 1,4,5-trisphosphate and diacylglycerol from PI45P2; by PLA2 to LPIs; PLA/lysophospholipases (LPLA1) to form the GPIs;and by PLD to form phosphatidic acid. PLC acts preferentially on PI45P2, whereas the other phospholipases may act on the different PIs (for simplicity in the figure,all the phospholipases are shown acting only on PI45P2).
Proposed functions of PX-domain proteins.a, Recruitment of the NADPH oxidase complex. Upon neutrophil activation, PI3K-I converts PI(4,5)P2 into PI(3,4,5)P3. The cytosolic subunits of the NADPH oxidase complex (p40, p47, p67) are recruited to developing phagosome at the plasma membrane by binding of the PX domain of p47phox to PI(3,4)P2 (green), generated upon dephosphorylation of PI(3,4,5)P3 by the 5-phosphatase SHIP-1. PI(3,4)P2 is then dephosphorylated by a PI(3,4)P2 4-phosphatase to generate PI(3)P (red), which binds to the PX domain of p40phox. The correct assembly of p40phox, p47phox and p67phox with the membrane-bound (cytb558) components of the complex results in a functional Phox complex that produces O2- b, Membrane trafficking. PI3K-II is recruited to the plasma membrane through binding of its PX domain to PI(4,5)P2and may promote the formation of clathrin-coated vesicles. Snx3 binds to PI(3)P (red) in the sorting early endosome and augments transport of transferrin (Tf) from the sorting to the recycling endosome. The yeast SNARE Vam7p is recruited by PI(3)P on multivesicular bodies (MVBs) and vacuoles to complex with other SNAREs and thereby promote vacuolar membrane docking and fusion.
O CH2 P OH O OH HC OC OC R1 R2 O O CH2 Simplified overview of the main synthetic pathways involved in the formation of polyphosphoinositides in higher plant cells Phosphatidic Acid serves as the precursor from which many of these second messenger lipids are derived → The two kinases, phosphatidylinositol 3-kinase (PtdIns 3K) and PtdIns(3)phosphate [PtdIns(3)P] 5-kinase (Fab1) are shown. Routes of synthesis that are established are shown by unbroken arrows, whereas steps that still need confirmation or are less well defined in vivo are indicated by broken arrows. Abbreviations: PtdIns(4)P, PtdIns(4)phosphate; PtdIns(5)P, PtdIns(5)phosphate; PtdIns(4,5)P2, PtdIns(4,5)bisphosphate; PtdIns(3,4)P2, PtdIns(3,4)bisphosphate; PtdIns(3,5)P2, PtdIns(3,5)bisphosphate
Lipid substrates and messengers produced by phospholipids- and/or galactolipid-hydrolyzing enzymes, and their downstream physiological effects Note that the substrate lipids can be located on the plasma membrane or other membranes, depending on the nature of a specific enzyme and its intracellular location
Phosphoinositides involved in classical phagocytosis PtdIns(4,5)P2 accumulates in pseudopods during extension of the phagocytic cup. As the phagosome seals, PtdIns(4,5)P2 disappears. This could be explained in part by its catabolism by phospholipases but also by its conversion into PtdIns(3,4,5)P3, and indeed, PtdIns(3,4,5)P3 appearance coincides with PtdIns(4,5)P2 clearance. PtdIns(3,4,5)P3 accumulates transiently in the phagocytic cup and is required for its closure. Once the phagosome is formed, PtdIns(3)P is produced on its surface and recruits proteins that control phagosome fusion and maturation. Other phosphoinositide species are present in the trans-Golgi complex (PtdIns(4)P) or in the nucleus (PtdIns(5)P), leading to the proposal that membrane identity can be mediated by compartmentalization of specific phosphoinositides
Phospholipid signaling under salt stress, drought, cold, or ABA. Osmotic stress, cold, and ABA activate several types of phospholipases that cleave phospholipids to generate lipid messengers (e.g., PA, DAG, and IP3), which regulate stress tolerance partly through modulation of gene expression. FRY1 (a 1-phosphatase) and 5-phosphatase-mediated IP3 degradation attenuates the stress gene regulation by helping to control cellular IP3 levels.
PLD and PA in response to H2O2PLD , is activated in response to H2O2 and the resulting PA functions in amplification of H2O2 -promoting PCD Stress stimulates production of H2O2 that activates PLD associated with the plasma membrane. Potential activators: Ca2+ and oleic acid. This increases PLD affinity to its substrates, stimulating lipid hydrolysis and PA production. PA may bind to target proteins, such as Raf-like MAPKK, that contain a PA binding moti, leading to the activation of MAPK cascades. PA may also function by modulating membrane trafficking and remodeling. These interactions modulate the cell's ability to respond to oxidative stress and decrease cell death. Dashed lines - hypothetical interactions.
Knockout of PLD renders Arabidopsis plants more sensitive to the reactive oxygen species H2O2 and to stresses H2O2 activates PLD , and PLD -derived PA functions to decrease the promotion of cell death by H2O2. These results suggest that both PLD and its product PA play a positive role in signaling stress responses PLD and its derivative PA provide a link between phospholipid signaling and H2O2-promoted cell death. PLD and PA positively regulate plant cell survival and stress responses. PLD & PA
The role of PLD in vesicular trafficking & signal transduction A) PLD catalytic activity. In the first step of the reaction (left panel), PLD removes the head group of a structural phospholipid, such as PC, forming covalent bond with the resulting phosphatidyl moiety, the PLD-PA intermediate (middle panel). In the second step (right panel), PLD transfers the phosphatidyl moiety to a nucleophile. Under physiological conditions, this is water, representing the hydrolysis of PC to generate PA. Primary alcohols, such as 1-butanol, can also be used as acceptors, resulting in the formation of PBut, a reaction that is used to measure PLD activity in vivo and in vitro (3, 8, 32). (B) Cytokinesis in plant cells. (C) Model of PLD binding to microtubules and membranes. PLD binds vesicular and plasma membranes through its covalent PLD-PA intermediate (Fig. 1A, middle panel). (D) PLD's contribution in PA signaling. A summary of factors activating PLD in plants and the role of PA in signaling
Phospholipid signalling pathways that are involved in plant defence responses. PLA2 generates lyso-phospholipids (LPL) and FFAs that stimulate the plasma membrane H+-ATPase, and free fatty acids can be metabolised via octadecanoid pathway to JA. PLC hydrolyses PIP2 into IP3 and DAG. IP3 diffuses into the cytosol, where it could release Ca2+ from intracellular stores, or is metabolised further to IP6. DAG remains in the membrane to be phosphorylated by DGK to PA. Activation of PLD generates PA directly by hydrolysing structural phospholipids such as PC. PA can activate MAPK, CDPK, ion channels, and NADPH oxidase, all of which are involved in typical defence-related responses. PA signalling is attenuated by its conversion to DGPP by PA kinase. All lipids or their derivatives that are involved in signalling are shown in red. Solid arrows indicate metabolic conversion; dashed arrows indicate activation (directly or indirectly) of downstream targets.
PI metabolism in Arabidopsis The different steps in the synthesis of PIs and the lipid kinases catalyzing the different reactions are indicated. PtdIns(3,4,5)P3 is present in animal cells but has not been detected in plant tissues, so far. In animal cells, PtdIns(3,4)P2 can be generated from PtdIns4P by a PtdIns 3-kinase or by an as-yet-unidentified PIPkin from PtdIns3P. Plant cells do not contain any homolog of the heterodimeric inositol lipid 3-kinases that are able to phosphorylate PtdIns4P to PtdIns(3,4)P2 and PtdIns(4,5)P2 to PtdIns(3,4,5)P3. PtdIns(4,5)P2 can be synthesized by type I and type II PIPkins from PtdIns4P and PtdIns5P, respectively. On the basis of sequence comparison, plants cells do not possess type II PIPkins. PtdIns5P is present in plants, but an enzyme capable of producing it has not been identified.
PLD is involved in O2.- production in Arabidopsis PLD suppression decreases Phosphatidic acid (PA) production PA levels increase during various stress conditions. PA-stimulated production of superoxide in PLD -deficient and wt leaves Plant Physiology, 2004, Vol. 134, pp. 129 Plant Physiol. 126 (2001) 1449-1
PA specifically induces leaf cell death in Arabidopsis • WT plants were infiltrated with PA or PC and photographed 24 h after treatment with the lipids. Arrows indicate the area of liposome infiltration. • Leaves of WT plants were floated on phospholipid liposomes • Trypan blue staining was used to visualize dying cells in areas of turgor loss in PA-treated leaves. Leaves of WT plants were detached, floated on PA (left), or PC (right) suspensions for 2 h, and stained with Trypan blue
Phospholipase C Activity These phospholipases are involved in second messenger generation from membrane phosphoinositides Phosphatidylinositol –4,5-bisphosphate (PIP2) O PLA2 O Diacylglycerol (DAG) O O Phospholipase C (PLC) hydrolysis PIP2 to yield two second messengers PLC O- O P O O- O- P O O Inositol-1,4,5-triphosphate (IP3) O 5 PLD 1 OH 4 OH HO O P O- O N.B. Different phospholipid specificities (releases different PIs) O-
The receptor for inositol 1,4,5-triphosphate (IP 3 )is located on the tonoplast and ER membranes Conformational changes in this receptor transduce subsequent signaling. Certain ion channel receptors,including the IP3 receptor,are composed of four subunits. Each subunit contains four membrane-spanning domains (not shown). When IP3 binds to the receptor,conformational changes result in movement of two of the subunits.The distribution of positive and negative charges stabilizes the open conformation of the channel and allows the entry of Ca2+ into the cytoplasm.
Domain structures of PLDa, PLDb, and PLDg in Arabidopsis XX in the PLD C2 marks the loss of two acidic residues potentially involved in Ca2+ binding; XX in the PPI-binding motifs marks the loss of the number of basic residues potentially required for PPI binding.
Direct and derived products of PLD activation LysoPA and free fatty acid (FA) can be formed from PA by nonspecific acyl hydrolase or by PLA. PA is dephosphorylated to DAG by PA phosphatase. CDP-DAG is the precursor for the synthesis of PS, PI, and PG. XOH, Primary alcohol used for transphosphatidylation; Ptd, phosphatidyl; NAE, N-acylethanolamine.
PI3K, phosphatidylinositol 3-kinase; PKC, protein kinase C; ARF, ADP-ribosylation factor; LPA, lysophosphatidic acid; PLA, phospholipase A; PA, phosphatidic acid; DGK, diacylglycerol kinase; PAP, PA phosphohydrolase; PIP5K, phosphatidylinositol 4-phosphate 5-kinase; MAPK, mitogen-activated protein kinase; MEK, MAP kinase kinase; ERK, extracellular signal-regulated kinase; SPHK, sphingosine kinase; Edg, endothelial differentiation gene
b 900-1315 g 1220-1285 d 600-870 Classes of Phospholipase C Four main isoforms (+variants) Animals only G protein activated Two main isoforms Animals only Tyrosine kinase activated Four mammalian isoforms + four splice variants All non-animal PLCs are in this class Ca2+ activated?
Pleckstrin homology – phosphoinositide binding Pleckstrin homology – phosphoinositide binding Catalytic X and Y domains Catalytic X and Y domains SH2 – phosphotyrosine binding SH2 – phosphotyrosine binding EF hand-like Ca2+ binding? EF hand-like Ca2+ binding? The Structure of PLCg Phosphotyrosines C3 – part of catalytic domain? Phospholipid interaction? P P P SH3 – interaction with cytoskeleton?
EGF binds to receptor EGF PLCg hydrolyses PIP2 to yield IP3 and DAG DAG P IP3 P P P PLCg phosphorylated Receptor phosphorylated Activation of PLCg by EGFR complex PLCg PIP2 PLCg phosphorylation may release it from interaction with inhibitor Inactive PLCg
Inositol-1,4,5-triphosphate (IP3) Diacyleglycerol (DAG) O- O P O O- O- P O O O O O 5 O 1 OH 4 OH HO O O P O- O O- PIP2-derived Second Messengers Hydrophilic Hydrophobic Binds to receptor on ER Remains in plasmalemma IP3 Receptor is Ca2+ channel Activates Protein Kinase C (PKC) AMPLIFICATION – many IP3/DAG per bound ligand
Summary Phosphatidylinositol-specific PLC hydrolyses membrane PIP2 PLCg has domains that allow binding to phosphotyrosine (SH2) PLCg associates with activated receptor tyrosine kinases PLCg is activated by tyrosine phosphorylation IP3 – soluble, induces Ca2+ release DAG – hydrophobic, activates protein kinase C Loewen, et al (2004). Phospholipid Metabolism Regulated by a Transcription Factor Sensing Phosphatidic Acid. Science304, 1644-1647. Inositol-induced alteration in phospholipid synthesis.
O- O- O P O- O P O- O O O O O O O O O P P P O- O- O- P P P P O- O- O- O- O O O O O O O O- O- O- O- O- O- O- Phosphatidylinositol 3’-Kinase (PI3K) Activity O O O Headgroup of PIP2 O- P O O- P O O- P O OH OH O O O 5 5 5 OH OH OH PI4K PI5K 4 4 4 1 1 HO HO OH OH 1 HO OH OH PI3K PI3K PI3K PTEN PTEN O O O OH O- P O O- P O O- P O OH O O O 5 5 5 PI5Ptase OH OH OH 4 4 4 1 1 1 OH OH OH OH PI3K phosphorylates inositol on the 3 position PTEN dephosphorylates inositol on 3 position
p85 binding ras binding Catalytic SH2 SH3 p110-binding Proline-Rich Class I PI3K p110a, b , d kinases p85a p85b adapters p55a, g p50a CLASS IA p110g kinase p101 adapter CLASS IB
Subunit interaction p110a phosphorylation of p85a (S608) inhibits PI3K activity p110a phosphorylation of p85a (S608) inhibits PI3K activity Proline-rich repeats bind SH3 domains of e.g. src, fyn or lck Proline-rich repeats bind SH3 domains of e.g. src, fyn or lck Class I PI3K Regulation p110d autophosphorylation inhibits PI3K activity regulation by p21ras SH2 bind pY-X-X-M SH2 also binds PI(3,4,5)P3 this binding competes with pY binding Inter-SH binds PI(4)P and PI(4,5)P2
Classical cPKC a, b1, b2 and g Activated by DAG and Ca2+ Novel nPKC d, e, and Activated by DAG but do not require Ca2+ Atypical aPKC z, i and l Do not require DAG or Ca2+ Protein Kinase C (PKC) Three Classes All forms require phosphatidylserine (PS) for activity cPKC have two zinc finger domains C1 – binds PS and Ca2+ C2 – binds DAG
Proliferation MARCKS Protein MARCKS Phosphorylation VEGF PKC Substrates PKC phosphorylation sites – release from membrane P P P MARCKS Myristoylation site – membrane association Calmodulin (CAM) binding • Associated with • Decreased MARCKS- F actin association • Actin polymerisation • Decrease CAM-dependent mlc phosphorylation
Effects mediated by PKC Proliferation - insulin MAP Kinase Pathway – ras independent Differentiation – wnt pathway +Apoptosis – UV-B, neutrophils (PKCd activated by caspase 3) -Apoptosis – suppresses Fas-induced PCD (PKCa?) CellPolarity – atypical PKC and interacting protein Feedback Inhibition of IP3/Ca2+ ReceptorDownregulation - e.g. EGF Inhibition of PLCg - -ve feedback STAT inhibition – PKCd blocks STAT DNA association
Summary Three classes of PKC All require phosphatidylserine for activity Pre-activation of PKC requires PDK-1 phosphorylation Activation completed by DAG (except aPKC class) MARCKS – major substrate for PKC MARCKS role in proliferation and cell morphology PKCs many roles in proliferation,differentiation and death
Summary PI3 kinases phosphorylate phosphoinositides at position 3 PTEN dephosphorylates phosphoinositides at position 3 p110 contains catalytic activity p85 responsible for recruiting enzyme to RTK PI-3,4,5-P3 recruits PDK1, PDK2 and Akt PI-3,4,5-P3 recruits other proteins and regulates cytoskeleton and transport
Ligand-activated RTK P110 phosphorylates PIP2 PDK1 phosphorylates Akt T308 (activation loop) PI(3,4,5)P3 pS124 pS124 T308 pT308 p110 S473 pS473 pT450 pT450 p85 Kinase? PDK1 PDK2 Fully-activated Akt Pre-activation of Akt T450 phosphorylation p85 binds to activated RTK Pleckstrin homology domain S124 T308 S473 Akt T450 Kinase Domain PI3K and Akt Activation PDK2 phosphorylates Akt S473 PI(4,5)P2 Akt, PDK1 and PDK2 all bind PIP3 (plekstrin homology domains)
Other PI-3,4,5-P3 Functions Regulation of Vesicle Transport either… • Binding to FYVE domain proteins • Regulating small GTP-binding protein Arf Rearrangement of actin cytoskeleton (rac) Recruitment of Tyrosine kinases – PH domains in Btk Enhancement of PLCg – direct interaction