Mastering Gene Regulation in Eukaryotes
550 likes | 593 Vues
Explore gene regulation in eukaryotes through genetics, mutations, transcription initiation, polymerases, trans-acting proteins, chromatin structure, and more. Understand regulatory elements like promoters and enhancers, and how RNA processing influences gene expression. Advance your knowledge with case studies like sex determination in Drosophila.

Mastering Gene Regulation in Eukaryotes
E N D
Presentation Transcript
Outline of Chapter 17 • How we use genetics to study gene regulation • Using mutations to identify cis-acting elements and trans-acting proteins • How genes are regulated at the initiation of transcription • Three polymerases recognize three classes of promoters • Trans-acting proteins control class II promoters • Chromatin structure affects gene expression • Signal transduction systems • DNA methylation regulates gene expression • How genes are regulated after transcription • RNA splicing • RNA stability • mRNA editing • Translation • Posttranslational modification • A comprehensive example of sex determination in Drosophila
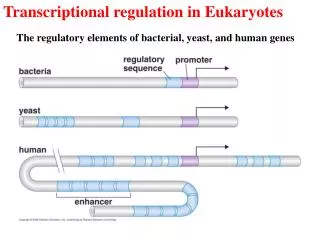
Regulatory elements that map near a gene are cis-acting DNA sequences • cis-acting elements • Promoter – very close to gene’s initiation site • Enhancer • can lie far way from gene • Can be reversed • Augment or repress basal levels of transcription Fig. 17.1 a
Reporter constructs are a tool for studying gene regulation • Sequence of DNA containing gene’s postulated regulatory region, but not coding region • Coding region replaced with easily identifiable product such as β-galactosidase (Lac Z) or green fluorescent protein (GFP) • Reporter constructs can help identify promoters and enhancers by using in vitro mutagenesis to systematically alter the presumptive regulatory region
Regulatory elements that map far from a gene are trans-acting DNA sequences because they encode transcription factors • Genes that encode proteins that interact directly or indirectly with target genes cis-acting elements • Known genetically as transcription factors • Identified by: • Mapping • Biochemical studies to identify proteins that bind in vitro to cis-acting elements Fig. 17.1 b
In eukaryotes three RNA polymerases transcribe different sets of genes • RNA polymerase I transcribes rRNA • rRNAs are made of tandem repeats on one or more chromosomes • RNA polymerase I transcribes one primary transcript which is broken down into 28S, 5.8S, and 18S by processing Fig. 17.2 a
RNA polymerase III transcribes tRNAs and other small RNAs (5S rRNA, snRNAs) Fig. 17.2 b
RNA polymerase II recognizes cis-acting regulatory regions composed of one promoter and one or more enhancers • Promoter contains initiation site and TATA box • Enhancers are distant from target gene • Sometimes called upstream activation sites Fig. 17.2 c
RNA polymerase II transcribes all protein coding genes • Primary transcripts are processed by splicing, a poly A tail is added to the 3’ end, and a 5’ GTP cap is added
Large enhancer region of Drosophila string gene • Fourteenth cell cycle of the fruit fly embryo • A variety of enhancer regions ensure that string is turned on at the right time in each mitotic domain and tissue type Fig. 17.3
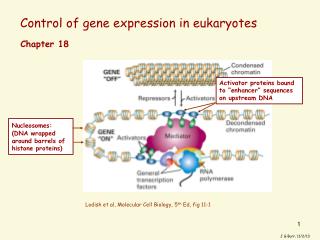
trans-acting proteins control transcription from class II promoters • Basal factors bind to the promoter • TBP – TATA box binding protein • TAF – TBP associated factors • RNA polymerase II binds to basal factors Fig. 17.4 a
Activator proteins • Also called transcription factors • Bind to enhancer DNA in specific ways • Interact with other proteins to activate and increase transcription as much as 100-fold above basal levels • Two structural domains mediate these functions • DNA-binding domain • Transcription-activator domain
Transcriptional activators bind to specific enhancers at specific times to increase transcriptional levels Fig. 17.5 a
Examples of common transcription factors • zinc-finger proteins and helix-loop-helix proteins bind to the DNA binding domains of enhancer elements Fig. 17.5 b
Some proteins affect transcription with out binding to DNA • Coactivator – binds to and affects activator protein which binds to DNA • Enhancerosome – multimeric complex of proteins • Activators • Coactivators • Repressors • Corepressors
Localization of activator domains using recombinant DNA constructs • Fusion constructs from three parts of gene encoding an activator protein • Reporter gene can only be transcribed if activator domain is present in the fusion construct • Part B contains activation domain, but not part A or C Fig. 17.6
Most eukaryotic activators must form dimers to function • Eukaryotic transcription factor protein structure • Homomers – multimeric proteins composed of identical subunits • Heteromers – multimeric proteins composed of nonidentical subunits Fig. 17.7 a
Leucine zipper – a common activator protein with dimerization domains Fig. 17.7 b
Repressors • Reduction of transcriptional activation but do not affect basal level of transcription • Activator-repressor competition • Quenching (corepressors) • Some repressors stop basal level of transcription • Binding directly to promoter • Bind to DNA sequences farther from promoter, contact basal factor complex at promoter by bending DNA causing a loop where RNA polymerase can not access the promoter
Transcription factors may act as activators or repressors or have no affect • Action of transcription factor depends on • Cell type • Gene it is regulating
Specificity of transcription factor can be altered by other molecules in cell • yeast a2 repressor – determines mating type • Haploid – a2 factor silences the set of “a” genes • Diploid – a2 factor dimerizes with a1 factor and silences haploid-specific genes Fig. 17.9
Myc-Max system is a regulatory mechanism for switching between activation and repression • Myc polypeptide has an activation domain • Max polypeptide does not have an activation domain Fig. 17.10
Myc-Max system is a regulatory mechanism for switching between activation and repression • As soon as a cell expresses the myc gene, the Max-Max homodimers convert to Myc-Max heterodimers that bind to the enhancers • Induction of genes required for cell proliferation Fig. 17.10
Gene repression results only when the Max polypeptide is made in the cell max gene Fig. 17.10 b
Gene activation occurs when both Myc and Max are made in cell Fig. 17.10
The locus control region is a cis-acting regulatory sequence that operates sequentially • Human b-globin gene cluster contains five genes that can all be regulated by a distant LCR (locus control region) Fig. 17.12 a
Proof that cis-acting factor such as LCR is needed for activation of b-globin gene Fig. 17.12 b
One mechanism of activation that brings LCR into contact with distant globin genes may be DNA looping Fig. 17.12 c
Other mechanisms of gene regulation • Chromatin structure • Slows transcription • Hypercondensation stops transcription • Genomic imprinting • Silences transcription selectively if inherited from one parent • Some genes are regulated after transcription • RNA splicing can regulate expression • RNA stability controls amount of gene product • mRNA editing can affect biological properties of protein • Noncoding sequences in mRNA can modulate translation • Protein modification after translation can control gene function
Remodeling of chromatin mediates the activation of transcription Fig. 17.13
Hypercondensation over chromatin domains causes transcriptional silencing. This is achieved by the methylation of cytosine residues Fig. 17.14
In mammals hypercondensation is often associated with methylation • It is possible to determine the methylation state of DNA using restriction enzymes that recognize the same sequence, but are differentially sensitive to methylation Fig. 17.14
Genomic imprinting results from chromosomal events that selectively silence genes inherited from one parent • 1980s, in vitro fertilization experiments in mice demonstrated pronuclei from two females could not produce a viable embryos
Experiments with transmission of Ig f2 deletion showed mice inheriting deletion from male were small. Mice inheriting deletion from female were normal. Figure 15.15 a
H19 promoter is methylated during spermatogenesis and thus the H19 promoter is not available to the enhancer and is not expressed
Epigenetic effect – whatever silences the maternal or paternal gene is not encoded in the DNA. The factor is outside the gene, but is heritable • Methylation can be maintained across generations by methylases that recognize methyl groups on one strand and respond by methylating the opposite strand Fig. 15.15 c
RNA splicing helps regulate gene expression Fig. 17.16
RNA stability provides a mechanism for controlling the amount of gene product • Cellular enzymes slowly shorten the poly-A tail. mRNA then degrades. • Length of poly-A tails of mRNAs affects the speed at which mRNAs are degraded after they leave the nucleus. • Histone transcripts receive no poly-A tail • mRNA quickly degrades after S phase of cell cycle
Specialized example of regulation through RNA stability Note also the untranslated sequences that help modulate their translation Fig. 17.17
mRNA editing can regulate the function of protein products – e.g., AMPA receptor gene in mammals Fig. 17.18
Protein modifications after translation provide a final level of control over gene function • Ubiquitination targets proteins for degredation • Ubiquitin – small, highly conserved protein. • Covalently attaches to other proteins • Ubiquitinized proteins are marked for degredation by proteosomes Fig. 17.19 a
Sex determination in DrosophilaA comprehensive example of gene regulation Sex specific traits in Drosophila Fig. 17.20
The X:A ratio regulates expression of the Sex lethal (sxl) gene • Key factors of sex determination • Helix-loop-helix proteins encoded by genes on the autosomes • Denominator elements • Helix-loop-helix proteins encoded by genes on the X chromosome • Numerator elements – monitor the X:A ratio through formation of homodimers or heterodimers • Sisterless-A and sisterless-B
Hypothesis to explain why flies with more numerator homodimers transcribe Sxl early in development • Numerator subunit homodimers may function as transcription factors that turn on Sxl • Females • Some numerator subunits remain unbound by denominator elements • Free numerator elements act as transcription factors at Pe promoter early in development • Males • Carry half as many X-encoded numerator subunits • All numerator proteins are bound by abundant denominator elements • Pe promoter is not turned on • The Sxl protein expressed early in development in females regulates its own later expression through RNA splicing • Females • Sxl protein produced early in development catalyzes the synthesis of more of itself through RNA splicing of the PL transcript • Males • No Sxl transcript in early development results in a unproductive transcript in later development from the PL promoter with a stop codon near the beginning of the transcript