Download

1 / 53
540 likes | 671 Vues
The role of acidity in tumours invasion. Antonio Fasano Dipartimento di Matematica U. Dini Firenze. IASI, Roma 11.02.2009. fasano@math.unifi.it. Invasion is not just growth. We will not deal with models describing just growth. Most recent survey paper:.

E N D
The role of acidity in tumours invasion Antonio Fasano Dipartimento di Matematica U. Dini Firenze IASI, Roma 11.02.2009 fasano@math.unifi.it
Invasion is not just growth We will not deal with models describing just growth Most recent survey paper: N. Bellomo, N.K. Li, P.K. Maini, On the foundations of cancer modelling: selected topics, speculations, & perspectives, Math. Mod. Meth. Appl. S. 18, 593-646 (2008)
There are several mechanisms of tumour invasion • intra and extra-vasation metastasis • enzymatic lysis of the ECM + haptotaxis • aggression of the host tissue by increasing acidity
Increase of acidity is originated by the switch to glycolytic metabolism Invasive tumours exploit a Darwinian selection mechanism through mutations The winning phenotypes may exhibit less adhesion increased mobility anaerobic metabolism (favoured by hypoxic conditions) higher proliferation rate
Anaerobic vs. aerobic metabolism ATP = adenosine triphosphate. Associated to the “energy level” Anaerobic metabolism Glycolytic pathway acid ( 2 ATP) Aerobic metabolism KREBS cycle Much more efficient in producing ATP Requireshigh oxygen consumption
The role of ATP production rate in the onset of necrosis in multicellular spheroids has been investigated only very recently • Most recent paper: • Bertuzzi, A.F., A. Gandolfi, C. Sinisgalli • Necrotic core in EMT6/Ro tumour spheroids: is it caused by an ATP deficit ? (submitted, 2009)
The level of lactate determines (through a complex mechanism) the local value of pH : lactate − + H + Cells in the glycolitic regime may increase their glucose uptake, thus producing more lactate As early as 1930 it was observed that invasive tumours (may) switch to glycolytic metabolism (Warburg)
The prevailing phenotype is acid resistant thanks to compensation mechanisms keeping the internal pH at normal levels Apoptosis threshold for normal cells: pH=7.1 (Casciari et al., 1992) for tumour cells: pH=6.8 (Dairkee et al., 1995) And the result is …
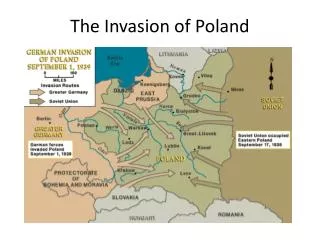
K. Smallbone, R.A.Gatenby, R.J.Gilles, Ph.K.Maini,D.J.Gavaghan. Metabolic changes during carcinogenesis: Potential impact on invasiveness. J. Theor. Biol, 244 (2007) 703-713. tumour host tissue H+ ions invasion front + GAP (from R.A.Gatenby-E.T.Gawlinski, 1996)
The model by Gatenby and Gawlinski consists in a set of equations admitting a solution chacterized by a propagating front with the possible occurrence of a gap Remark: as it often happens for mathematical models of tumours, it must be taken with some reservation !
The G.G. model One space dimension s = time logistic damage Host tissue tumour H+ ions reduced diffusivity large production decay small Defects: mass conservation? Damage on the tumour? Metabolism? Diffusion as main transport mechanism?…
Non-dimensional variables prol. rate host tissue decay/production carrying capacities ions diffusivity damage rate very small Basic non-dimensional parameters
The normalized G.G. model One space dimension (space coord.x) Normalized non-dimensional variables: all concentrations vary between 0 and 1 Normalized logistic growth rate Acidic aggression a>0 Host tissue d<<1, b>1 tumour c>0 H+ ions production - decay Normalized diffusivity
Search for a travelling wave Set Z = x t with Solutions of this form, plotted vs. x, are graphs which, as time varies, travel with the speed || to the right (>0: our case), or to the left (<0)
The system becomes etc.
Conditions at infinity corresponding to invasion Normal cells: max(0,1a) 1 Tumour cells: 1 0 H+ ions : 1 0 For a<1 a fraction of normal cells survives A. Fasano, M.A. Herrero, M. Rocha Rodrigo: study of all possible travelling waves (2008), To appear on Math. Biosci.
REACTION - DIFFUSION and travelling waves M.A. Herrero : Reaction-diffusion systems: a mathematical biology approach. In Cancer modelling and simulation, L. Preziosi .ed , Chapman and Hall ( 2003), 367-420.
The prototype: Fisher, 1930 Kolmogorov-Petrovsky-Piskunov, 1937 (spread of an advantageous gene) F slope=a x 0<u<1 for t>0 by the max principle 1 0 The solution is asymptotic to a TRAVELLING WAVE with speed
Existence of travelling waves Fisher’s equation 1 c 0 Existence requires: Describes an invasive process
There are two equilibrium points: U=0, U=1 The eigenvalues associated to linearized near U=0 are the solutions of and are both real and positive if so thatthe equilibrium is unstable Remark: complex eigenvalues produce oscillations (= sign changes !)
Taking V=1U and linearizing near V=0 theequation for the eigenvalues becomes Now the eigenvalues are real but with opposite sign (U=1 is a saddle point) A wave has to take from the unstable to the stable equilibrium
In the phase plane p=U’, U p connecting heteroclinic 1 0 U unstable node saddle The connecting heteroclinic corresponds to the travelling wave The solutions of the original p.d.e.’s system with suitable initial data converge to the slowest wave for large time
For their system Gatenby and Gawlinski computed just one wave with speed proportional to Good guess ! This is conjectured to describe the large time asymptotic behaviour of the solution of the initial-boundary value problem for the original p.d.e.’s system The conjecture is based on the similarity to the famous two-population case The proof is still missing (due to higher dimensionality)
Two classes of waves: • slow waves: = 0d(d<<1): singular perturbation !!! • fast waves: = O(1) as d 0 Difficult but physical ! A mathematical curiosity ? Slow waves: Technique: matching inner and outer solutions Take = z/d as a fast variable:look at the front region with a magnifying lens
For all classes of waves u can be found in terms of w w can be found in terms of v
The equation is of Bernoulli type The equation has the integral Asymptotic convergence rate determined by wave speed
Summary of the results slow waves: = 0d 0 < ½, No solutions for >½ similar to Fisher’s case The parameter adecides whether the two cellular species overlap or are separated by a gap
Normal cells extends to 0 < a 1 0 1 < a 2 overlapping zone Thickness of overlapping zone
a > 2 0 gap Thickness of gap
For any a > 0 tumour limit case Fsolution of the Fisher’s equation This is the slowest possible wave H+ ions
Numerical simulations = ½ , minimal speed The propagating front of the tumour is very steep as a consequence of d<<1 (this is the case treated by G.G.)
0 < a 1 Survival of host tissue
1 < a 2 Overlapping zone
a > 2 gap
Remarks on the parameters used by G.G. Using the data of Gatenby-Gawlinski the resulting gap is too large Possible motivation: make it visible in the simulations Reducing the parameter a from 12.5 (G.G.) to 3 produces the expected value (order of a few cell diameters)
a = 3 (keeping the same scale) b = 10 b = 1 (G.G.) The value of b only affects the shape of the front b = ratio of growth rates, expected to be>1
Fast waves ( = O(1)) No restrictions on > 0
Linear stability of fast waves Let Then the system has solutions of the form for a 1
Other invasion models are based on a combined mechanism of ECM lysis and haptotaxis (still based on the analysis of travelling waves)
[ICM Warsaw] J.Math.Biol., to appear HSP’s increase cells mobility
Analysis of multicellular spheroids in a host tissue Folkman-Hochberg (1973) Acid-mediated invasion
Acid is produced in the viable rim and possibly generates a gapand/or a necrotic core host tissue Necrotic core Viable rim host tissue gap Evolution described as a quasi-steady process
K. Smallbone, D. J. Gavaghan, R. A. Gatenby, and P. K. Maini. The role of acidity in solid tumour growth and invasion. J. Theor. Biol. 235 (2005), pp. 476–484. Vascular and avascular case, gap always vascular, no nutrient dynamics(H+ ions produced at constant rate by tumour cells) L. Bianchini, A. Fasano. A model combining acid-mediated tumour invasion and nutrient dynamics, to appear on Nonlinear Analysis: Real World Appl. (2008) Vascularization in the gap affected by acid, acid production controlled by the dynamics of glucose Many possible cases / Qualitative differences
Two cases: avascular, vascular Host tissue sorrounding tumour Glucose concentration gap P Q H+ ions concentration h n.c. The viable rim is divided in a proliferating and in a quiescent region: >P , <P viable rim gap For the vascular case we use a factor (h) reducing the vascular efficiency possibly to zero All boundaries are unknown !
Necrotic core Host tissue (vascular) gap 0 RH r1 rP r2 r3 spheroid radius quiescent proliferating The necrotic core, the gap and the quiescent regionmay or may not exist, depending on the spheroid size r2 The necrotic core may have different origin many different combinations are possibleboth in the vascular and in the avascular case
Necrotic core Host tissue (vascular) quiescent Prol. gap RH * and h flat r1 rP r2 r3 = P = glucose concentration: diffusion-reaction h = H+ ions concentration: product of metabolism Concentrations and fluxes continuous at interfaces Diffusion of glucose and of H+ ions is quasi-steady
Necrotic core Host tissue (vascular) quiescent Prol. gap RH * and h flat r1 rP r2 r3 = P • STRATEGY: • suppose r2is known and compute all other quantities • consider some (naive) model for the growth of the spheroid and deduce the value of r2at equilibrium By increasing r2all combinations are encountered
The avascular case prol. threshold death threshold production rate removal rate by vasculature consumption rate
Necrotic core Host tissue (vascular) quiescent Prol. gap h 0 1 and h flat RH r1 rP r2 r3 = P Conditions are found onr2for the onset of interfaces