Download

1 / 30
330 likes | 625 Vues
Lecture 6 Import to mitochondria , chloroplasts, peroxisomes and nuclei. We shall see that 1) Import into these organelles is mainly or wholly post-translation 2) In all cases except for peroxisomes the proteins must be routed to the right sub-compartment

E N D
Lecture 6 Import to mitochondria, chloroplasts, peroxisomes and nuclei We shall see that 1) Import into these organelles is mainly or wholly post-translation 2) In all cases except for peroxisomes the proteins must be routed to the right sub-compartment 3) The structures controlling import are simple in peroxisomes, complex in mitochondria and chloroplasts and very sophisticated in the case of the nuclear pore
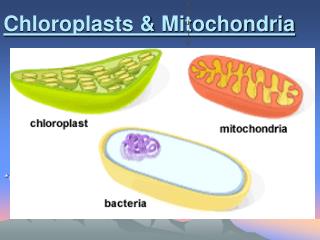
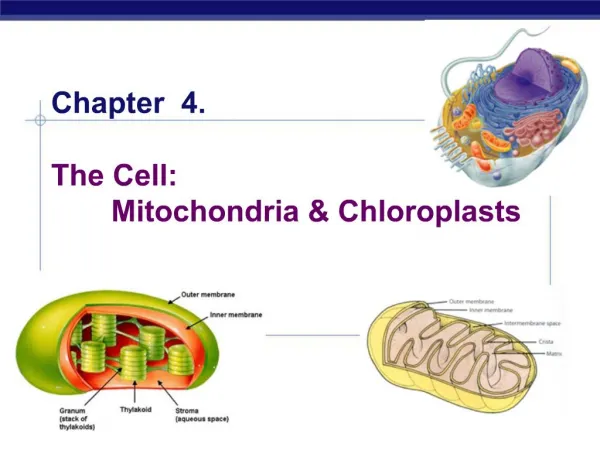
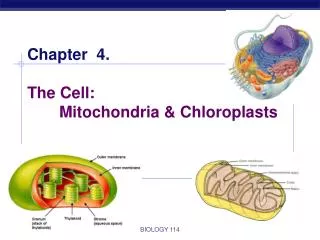
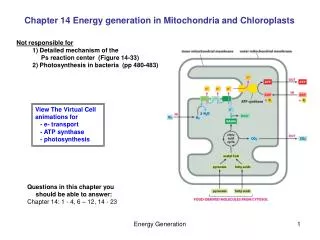
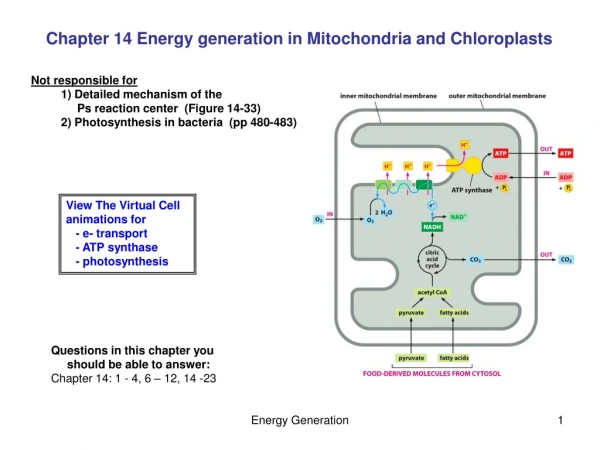
Mitochondrial and chloroplast subcompartments 1) The outer membrane 2) The inter membrane space 3) The inner membrane 4) The matrix and in chloroplasts also 5) The thylakoid membrane 6) The lumen of the thylakoid
Recognition of proteins destined for mitochondria • This involves both terminal signal sequences and internal signal patches • In some cases proteins for import are packaged with chaperones, especially Hsp70, but in other cases it would appear that the fully-folded protein is imported intact. • There are two major recognition sites on the outer membrane, one recognising fully folded proteins, the other proteins associated with chaperones
Targetting to mitochondria Targetting of proteins to mitochondria involves both internal and terminal signal sequences and signal patches
Transfer across the Outer membrane • This involves the TOM (Transport - Outer Membrane) complex which is comprised of at least 8 proteins • There appear to be three methods for recognising proteins destined for import. • Some proteins bind to a TOM20/22 which in turn binds to the universal transporter TOM40 • Others appear require binding to an inner membrane protein OXA1-details under dispute • Finally yet others bind to TOM70 which presents them to TOM40
Continued • Transfer across the outer membrane does not require ATP hydrolysis directly but obviously energy is needed as the proteins are moving up a concentration gradient. The energy is probably provided by ATP hydrolysis in binding and releasing Hsp70 • The transfer of proteins destined for the matrix is halted. This may occur soon after the outer membrane is contacted (the so called cis site) but the more important trans site halts translation leaving a length of amino acids projecting into the intermembrane space
The Transfer - Inner membrane complex • This again is made up of a number of peptides. This complex performs a number of functions 1) It must recognise the signal on peptides projecting from the TOM complex 2) It must guide these through the inner membrane and then pull the rest of the molecule through. • The matrix targetting signal must now be removed. This is carried out by a special Matrix processing peptidase, sometimes assisted by a second enzyme, the mitochondrial intermediate peptidase
Contimued • Movement of proteins through the pore depends on the maintenance of the potential difference across the membrane. This is normally app. 200 mV which is equivalent to 400,000 V/cm • The key role in dragging the remainder of the proteins through the membrane is played by the mitochondrial form of Hsc70. Transfer requires ATP hydrolysis. Three mechanisms have been suggested. The ratchet mechanism proposes that transfer is by Brownian motion with Hsc70 binding preventing back movement. The molecular motor model proposes that the conformational change of mtHsc70 results in the protein being pulled across the membrane
TOMs and TIMs • It is clear that there is no permanent conncetion between the two complexes. It is less clear whether there are temporary around the transfer pore connections or whether the sole junction is by the protein in transit. The inner membrane protein TIM54 has a projection which may reach the inner membrane • It is unclear whether there are permanent contact sites between the inner and outer membrane or whether in real life the outer membranes lie close to the outermost parts of the inner membrane
Further processing • Proteins are transferred across the TOM/TIM system as extended chains. A variety of chaperone proteins assist in the correct folding following transfer to the matrix • Chaperones also assist in the assembly of inner membrane components which contain both polypeptides specified by nuclear genes and polypeptides specified by mitochondrial DNA
Other pathways • Little is known about the insertion of outer membrane proteins. The most abundant protein is porin which cinsists almost entirely of transmembrane beta sheets • Apocytochrome C appears to transverse the outer membrane by a special reversible pathway. Conversion to cytocrome C makes the process irreversible. • Lactate dehydrogenase has two targetting sequnces - a matrix targetting sequence and a sequence which specifies the intermembrane space. It is not clear whether there is sequential transfer or the second signal acts as a “stop transfer”
Chloroplasts • Transfer in chloroplasts is very similar to transfer in mitochondia. Thylakoid proteins carry two signal sequences, one specifies matrix, the second thylakoid
Import of Proteins into Peroxisomes • Peroxisomes are a type of microbody. Microbodies are cell organelles bounded by a single membrane and are used for a variety if different processes. For example peroxisomes contain enzymes which produce hydrogen peroxide (and have the means for destroying it). In addition plants have glyoxysomes which contain the enzymes of the glyoxylate cycle and yeasts have a variety of microbodies including ones involved in methanol oxidation.
Targeting to peroxisomes • Fortunately this is nice and simple. Two major targetting signals Pex5 (PTS1R) and PTS2R have been identified. The Pex5 signal is a carboxy-terminal tripeptide SKL. • The mechanism of Prx5appears to involve binding of the SKL sequence to Pex5. This then interacts with a peroxisomal membrane protein called Pex14 forming a channel. It is not clear whether Pex5 and the protein move together across the channel or whether the imported protein is “pushed” through. • In the peroxisome the signal sequences are removed and Pex5 is recycled with the help of Pex2, 10, and 12.
To be noted • ATP hydrolysis is required for import • Import into peroxisomes does not require unfolding of the protein chain - even gold particles conjugated to a peroxisomal protein are imported. • Hsp70 is however needed and becomes bound to the exterior of the peroxisome. • .Study of patients with Zellweger’s syndrome, where import of proteins into peroxisomes shows firstly that the peroxisomal membrane proteins are incorporated by another route and secondly that at least 8 polypeptides, including Pex2, are involved in the transfer.
Peroxisome matrix and membrane proteins enter by different routes
The Nucleus • The outer nuclear membrane is continuous with the ER but the inner nuclear membrane, the nuclear lamina and the chromatin need to be supplied with proteins. The nuclear pore complex is a sophisticated gateway
The nuclear pore complex • The nuclear pore complex allows large complexes such as ribosomes to pass through while retaining small peptides within the nucleus • The complex may consist of up to 100 polypeptide chains • The complex consists of two rings of particles with 8 spokes extending towards a central 26 nm pore
Import of Proteins into the nucleus • This is specified by a number of signal sequences most of which are internal • These signals are recognised by families of cytosolic receptors. These may act as chaperones. • A small GTP-binding protein called ran plays a central role • As with other small GTP-binding proteins a key role is played by the GTP/GDP exchange factor (GEF) and the GTPase activiating protein (GAP). Ran binding protein assists in GTP hydrolysis
The mechanics of import • The protein to be transported binds to an import receptor and moves in to the nucleus. • Ran-GTP binds to the import receptor releasing the cargo • The receptor:Ran:GTP complex moves out of the nucleus • The GTP is hydrolysed • Ran:GDP is released and returns to the nucleus leaving the receptor free to bind another protein • Ran GEF catalyses then exchange of GTP for GDP
And of export • The empty export receptor enters the nucleus and binds the cargo protein and Ran:GTP. • This passes out of the nucleus • In the cytoplasm GTP is hydrolysed and GDP:Ran and the cargo protein are released • GPD:Ran and the esport receptor return separately to the nucleus. • The GDP bound to Ran is exchanged for a GTP
References • The basic pathways for import of proteins into mitochondria have been understood for some time but the details keep changing. Several of the diagrams come from Lodish 5th Edn where the pictures are on the web. In www.whfreeman.com • More on Ran may be found in Azuma and Dasso, COCB 12 (2000)301-307 • Ryan and Wente’s review of the “Nuclear pore complex …) COCB 12 (2000) 361-371is almost unreadable but is well referenced