Download

1 / 49
500 likes | 1.04k Vues
Chapter 8. Energy Generation in Mitochondria and Chloroplasts. (1) Mitochondria: in all eukaryotic cells The relationship between the structure and function of mit. (2) Chloroplasts: in plant cells The relationship between the structure and function of chl.

E N D
Chapter 8 Energy Generation in Mitochondria and Chloroplasts (1) Mitochondria: in all eukaryotic cells The relationship between the structure and function of mit. (2) Chloroplasts: in plant cells The relationship between the structure and function of chl. Mit: Oxidative phosphorylation → ATP Chl: Photosynthesis → ATP + NADPH → Sugar
1. Mitochondria and oxidative phosphorylation • The size and number of mitochondria reflect the energy requirements of the cell. A. Mitochondrial structure and function Figure7-4Relationship between mitochondria and microtubules.
Figure7-3Mitochondrial plasticity.Rapid changes of shape are observed when a mitochondrion is visualized in a living cell.
Figure7-5Localization of mitochondria near sites of high ATP utilization in cardiac muscle and a sperm tail.
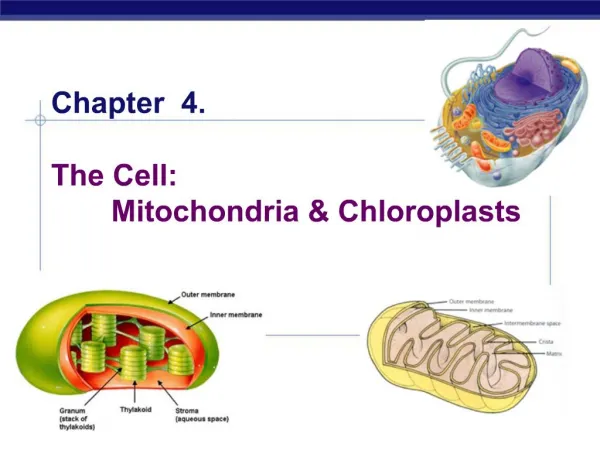
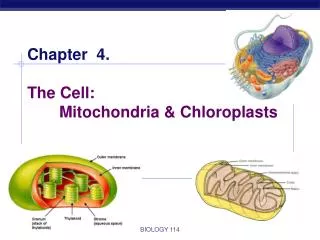
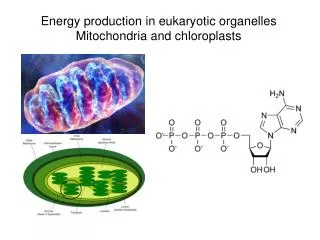
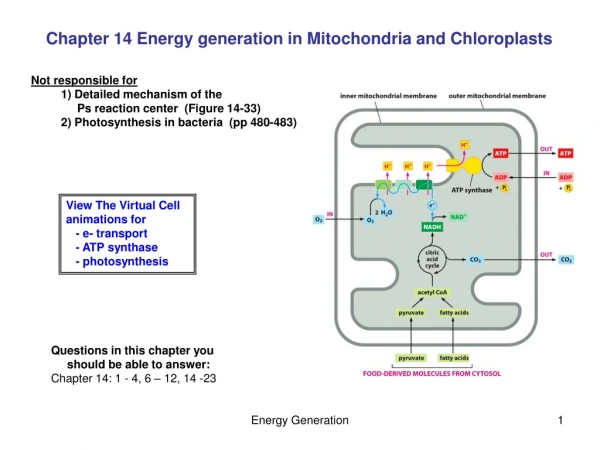
Inner and outer mitochondrial membranes enclose two spaces: the matrix and intermembrane space.
Outer membrane: Contains channel-forming protein, called Porin. Permeable to all molecules of 5000 daltons or less. Inner membrane (Impermeability): Contains proteins with three types of functions: (1) Electron-transport chain: Carry out oxidation reactions; (2) ATP synthase: Makes ATP in the matrix; (3) Transport proteins: Allow the passage of metabolites Intermembrane space: Contains several enzymes use ATP to phosphorylate other nucleotides. Matrix: Enzymes; Mit DNA, Ribosomes, etc.
B. Specific functions localized within the Mit by disruption of the organelle and fractionation Figure14-6Fractionation of purified mitochondria into separate components.These techniques have made it possible to study the different proteins in each mitochondrial compartment. The method shown, which allows the processing of large numbers of mitochondria at the same time, takes advantage of the fact that in media of low osmotic strength water flows into mitochondria and greatly expands the matrix space (yellow). While the cristae of the inner membrane allow it to unfold to accommodate the expansion, the outer membranewhich has no folds to begin withbreaks, releasing a structure composed of only the inner membrane and the matrix.
Inner membrane: Outer membrane: Phospholipid synthesis Electron transport Localization of metabolic functions within the mitochondrion fatty acid desaturation Oxidative phosphorylation Fatty acid elongation Metabolite transport Matrix Intermembrane space Pyruvate oxidation Nucleotide phosphorylation TCA cycle ß oxidation of fats DNA replication, RNA transcription, Protein translation
A. Molecular basis of oxidation: Electron- transport chain
The structure of the ATP synthase B. Molecular basis of phosphorylation: ATP synthase F1 particle is the catalytic subunit; The F0 particle attaches to F1 and is embedded in the inner membrane. F1: 5 subunits in the ratio 3:3:1:1:1 F0: 1a:2b:12c
Proton translocation through F0 drives ATP synthesis by F1: Binding Change Model and rotational catalysis Boyer proposed in 1979, and was greatly stimulated by the publication in 1994 of the structure for F1 complex (X-ray) from bovine heart mitochondria
Direct experimental evidence supporting the rotational catalysis. Japan researcher, Nature 386: 300, 1997.
Other roles for the proton-motive force in addition to ATP synthase
The pH and electrical gradient resulting from transport of protons links oxidation to phosphorylation. C. Mithchell’s Chemiosmotic theory (1961) • When electrons are passed to carriers only able to accept electrons, the H+ is translocated across the inner membrane. More than 21026 molecules (>160kg) of ATP per day in our bodies.
Electrons pass from NADH or FADH2 to O2, the terminal electron acceptor, through a chain of carriers in the inner membrane (FMN, Fe-S center, Heme group Fe, CoQ); As electrons move through the electron-transport chain, H+ are pumped out across the inner membrane, and form Proton motive force; Electrons move through the inner membrane via a series of carriers of decreasing redox potential
Figure7-26An experiment demonstrating that the ATP synthase is driven by proton flow.By combining a light-driven bacterial proton pump (bacteriorhodopsin), an ATP synthase purified from ox heart mitochondria, and phospholipids, vesicles were produced that synthesized ATP in response to light. If not all the detergent is removed, what will happen?
Summary of the major activities during aerobic respiration in a mitochondrion NADHO2: 3ATP/2e; FADH2 O2 : 2ATP/2e
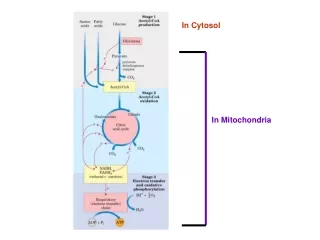
生物氧化产生ATP的统计 一个葡萄糖分子经过细胞呼吸全过程产生多少ATP? 糖酵解:底物水平磷酸化产生 4 ATP(细胞质) 己糖分子活化消耗 2 ATP(细胞质) 产生 2NADH,经电子传递产生 4或 6 ATP (线粒体)净积累 6或8 ATP丙酮酸氧化脱羧:产生 2NADH(线粒体),生成 6ATP三羧酸循环:底物水平的磷酸化产生(线粒体)2ATP;产生 6NADH(线粒体),生成18ATP;产生 2FADH2(线粒体),生成 4 ATP总计生成 36或38 ATP
3. Chloroplast and photosynthesis A. Comparison of a mitochondrion and a chloroplast.
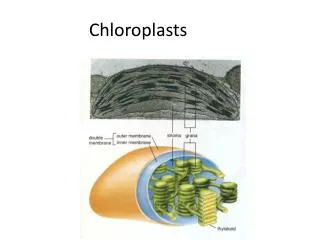
Figure14-39The chloroplast.This photosynthetic organelle contains three distinct membranes (the outer membrane, the inner membrane, and the thylakoid membrane) that define three separate internal compartments (the intermembrane space, the stroma, and the thylakoid space). The thylakoid membrane contains all of the energy-generating systems of the chloroplast. In electron micrographs this membrane appears to be broken up into separate units that enclose individual flattened vesicles (see Figure 14-40), but these are probably joined into a single, highly folded membrane in each chloroplast. As indicated, the individual thylakoids are interconnected, and they tend to stack to form aggregates called grana.
(A) A wheat leaf cell in which a thin rim of cytoplasm containing chloroplasts surrounds a large vacuole. (B) A thin section of a single chloroplast, showing the starch granules and lipid droplets that have accumulated in the stroma as a result of the biosyntheses occurring there. (C) A high-magnification view of a granum, showing its stacked thylakoid membrane. (Courtesy of K. Plaskitt.)
Figure7-42Photosynthesis in a chloroplast.Water is oxidized and oxygen is released in the photosynthetic electron-transfer reactions, while carbon dioxide is assimilated (fixed) to produce carbohydrate in the carbon-fixation reactions.
C. The antenna complex and photochemical reaction center in a photosystem
Light-dependent reaction: Electron transport in the thylakoid membrane and noncyclic photophosphorylation:
Carbon dioxide fixation and the synthesis of carbohydrate in C3 plants (Calvin cycle)
Figure14-43The initial reaction in carbon fixation.This reaction, in which carbon dioxide is converted into organic carbon, is catalyzed in the chloroplast stroma by the abundant enzyme ribulose bisphosphate carboxylase. The product, 3-phosphoglycerate, is also an important intermediate in glycolysis: the two carbon atoms shaded in blue are used to produce phosphoglycolate when the enzyme adds oxygen instead of CO2。
4. Organelle DNA and protein importing • The size range of organelle DNA is similar to that of viral DNAs. • Mit DNA: from <6,000bp (plasmodium falciparum) ~ >300,000bp (some land plants). DNA of Mit genome (in mammals) ≈16,500bp(<0.001% of nuclear genome) ; Chl genomes are about 10 times larger and contain about 120 genes. • Chl DNA: from 70,000to 200,000bp (genome of land plants); A. Organelle DNA
Genes in mtDNA encode rRNAs, tRNAs, and some mitochondrial proteins • Human mt DNA: 16,569bp • 2 rRNAs, 22 tRNAs, • 13 polypeptides: NADH reductase. 7 sub. • Cty b-c1 complex. 1 cytb • Cyt oxidase. 3 subunits • ATP synthase: 2 F0 sub Products of mt genes are not exported
B. Mit and Chl have their own genetic systems Mit and Chl are organelles semiautocephaly. The synthesis of mt proteins is coordinated
Tree proteins translocators in Mit membranes: • TOM, TIM,and OXA complex are multimeric membrane protein, that catalyze protein transport across Mit membrane, TOM, TIM stand for translocase of the outer and inner Mit membranes respectively. • TOM functions across the outer membrane; TIM(TIM23 and TIM22) function across the inner membrane. • OXA mediates the insertion of inner membrane proteins that are synthesized within the Mit. OXA also helps TOM and TIM to insert some proteins into the matrix. C. The transport protein into Mit. And Chl.
Translocation of precursors to the matrix occurs at the sites where the outer and inner membranes are close together;
The protein import by Mit: N-terminal signal sequence is recognized by receptors of TOM; The protein is translocated across both Mit membranes at or near special contact sites.
Only unfolded proteins can be imported into Mit; • Mit precursor proteins remain unfolded through interactions with hsp70 chaperone proteins in the cytosol after they are synthesized. • ATP hydrolysis and H+ gradient are used to dtive protein import into Mit
Protein transport into the inner Mit membrane and the intermembrane space requires two signal sequences
Two signal sequences are required to direct proteins to the Thylakoid membrane in Chl. Translocation into thylakoid space or thylakoid M can occur by any one of at least four routes.
5. The proliferation and origin of Mit and Chl. A.Organelle growth and division determine the number of Mitochondria and Plastids in a cell • Mit fission and fusion (a dividing Mit in a liver cell); Dividing or Budding of Mit. • Chloroplasts: dividing and formation of chloroplasts from proplastids begins by the light-induced budding of the inner membrane.
B. Origin: The endosymbiont theory • Compare the ribosomal RNA with the base sequence of various bacterial rRNAs: • Purple bactria-Mitochondria • Cyanobacteria-Chloroplasts