Download
1 / 194
1.98k likes | 2.36k Vues
第六章 RNA 转录 ( RNA transcription ). 6.1. 基本概念 6.2. 原核生物 RNA 转录的起始 6.3. 真核生物 RNA 转录的起始 6.4. 转录的延伸 6.5. 转录的终止 6.6. 原核生物转录产物的后加工 6.7. 真核生物转录产物的后加工 6.8. 真核生物转录产物中内元的去除 6.9. 不连续转录和反式拼接. 第一节 基本概念. 转录 (transcription): 是指以 DNA 为模板,在依赖于 DNA 的

E N D
第六章 RNA转录 (RNA transcription)
6.1. 基本概念 6.2. 原核生物RNA转录的起始 6.3. 真核生物RNA转录的起始 6.4. 转录的延伸 6.5. 转录的终止 6.6. 原核生物转录产物的后加工 6.7. 真核生物转录产物的后加工 6.8. 真核生物转录产物中内元的去除 6.9.不连续转录和反式拼接
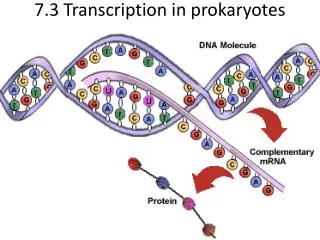
转录(transcription):是指以DNA为模板,在依赖于DNA的转录(transcription):是指以DNA为模板,在依赖于DNA的 RNA聚和酶催化下,以4中rNTP(ATP、 CTP、GTP和UTP)为原料,合成RNA 的过程。 在有些病毒中,RNA也可以指导合成RNA。 • 若 干 基 本 概 念 • 是基因表达的第一步,也是最关键的一步。 • 以Double Strand DNA中的一条单链作为转录模板 (杂交实验所证实)
有义链 (sense strand) • [又称编码链 coding • strand]: • 指不作模板的DNA单链 • 反义链 (antisense strand) • [又称模板链 template • srand] : 指作为模板进行RNA转录的链 (60年代以前的表示方法与此相反)
在依赖DNA的RNA聚合酶作用下进行转录 • A=U、C≡G 合成RNA分子 • 转录合成RNA链的方向为5’→3’,模板单链DNA的极性 方向为3’→5’, 而非模板单链的极性方向与RNA链相 同,均为5’→3’。(书写) • 基因转录方式为不对称转录(一条单链DNA 为模板,RNA 聚合酶的结合)
-10 +1 +10 upstream start point downstream • RNA 的转录包括 promotion. elongation. termination 三过程 • 从启动子 (promoter)到终止子(terminator)称为转录单 位 (transcription unit) (转录起始点) • 原核生物的转录单位多为 polycistron in operon,真 核生物中的转录单位多为monocistron, No operon • 转录起点即转录原点记为+1,其上游记为负值,下游 记为正值
第二节 原核生物RNA转录的起始(RNA Transcription promotion in prokaryots)
Absence of lactose z y a i p o Active No lac mRNA Presence of lactose z y a i p o Inactive -Galactosidase Permease Transacetylase 两个相关概念: * 操纵元(operon): 是原核生物基因 表达调控的一个 完整单元,其中 包括结构基因、 调节基因、操作 子和启动子
β α β α σ σ + α β’ α β’ Core Enzyme Holo Enzyme • 启动子(promoter):是指DNA分子上被RNA聚合酶识别 • 并结合形成起始转录复合物的区域 ,它还包括一些调节 • 蛋白因子的结合位点 (频率、效率) 一、原核生物的RNA polymerase (E. coli) 1、全酶 (Holo Enzyme)和核心酶(Core Enzyme)
(1) 全酶(Holo Enzyme) • 用于转录的起始 • 依靠空间结构与DNA模板结合 (σ与核心酶结合后 引起的构象变化) • 专一性地与DNA序列(启动子)结合 • 结合常数:1014/mol • 半衰期:数小时 (107/mol 1秒以下) • 转录效率低,速度缓慢( σ的结合)
(2) 核心酶 (Core Enzyme) • 作用于转录的延伸过程(终止) • 依靠静电引力与DNA模板结合(蛋白质中碱性 基团与DNA的磷酸根之 间) • 非专一性的结合(与DNA的序列无关) • 结合常数:1011/mol • 半衰期:60秒
(3)全酶的组装过程 2 α+ β α2 β+ β’ α2ββ’+σ α2ββ’σ 此外,新发现的一种ω亚基的功能尚不清楚。 2、各亚基的特点和功能 (1) σ因子 • σ因子可重复使用 • 修饰RNApol构型 • 使Holo Enzyme 识别启动子的Sextama Box(-35区), 并通过σ与模板链结合
• 不同的σ因子识别不同的启动子 E.coli 中有五种σ因子(σ70、σ32、σ54、σ28、 σ24) 枯草杆菌中有11种σ因子 (σ因子的更替对转录起始的调控) (2)α因子 • 核心酶的组建因子 α+α 2α+βα2β+β’ • 促使RNApol 与DNA模板链结合 • 前端α因子--使模板DNA双链解链为单链 • 尾端α因子--使解链的单链DNA重新聚合为双链
(3) β因子 • 促进RNApol+NTP RNA elongation • 完成NMP之间的磷酸酯键的连接 • Editing 功能(排斥与模板链不互补的碱基) • 与Rho (ρ)因子竞争RNA 3’-end
• 构成Holoenzyme 后,β因子含有两个位点 I site(initiation site . Rifs): 该位点专一性地结合 ATP或者GTP (需要高浓度的ATP或GTP) E site(elongation site. RifR):对NTP非专一性地结 合(催化作用和Editing功能) (4) β’ 因子 • 参与RNA非模板链(sense strand)的结合 (充当SSB)
由于各亚基的功能,使全酶本身含有五个功能位点由于各亚基的功能,使全酶本身含有五个功能位点 • 有义DNA链结合位点(β’亚基提供) • DNA/RNA杂交链结合位点(β亚基提供) • 双链DNA解链位点(前端 α亚基提供) • 单链DNA重旋位点(后端α亚基提供) • σ因子作用位点
E. coli RNA polymerase 155 KD 36.5 KD 11 KD 36.5 KD 70 KD 只用于起始 151 KD 用于起始和延伸
二、启动子(promoter)的结构与功能 (Prok.E.coli) 1、启动子由两个部分组成 (1) 上游部分— CAP-cAMP结合位点 (基因表达调控的正控制位点) CAP(catabolite gene Activator Protein) 降解物基因活化蛋白,环腺苷酸(cAMP)的受体蛋白 (2) 下游部分— RNApol的进入(结合)位点 -35 ~ -10 包括识别位点和结合位点(R B位点)
2、 RNA聚合酶的进入位点 (1) Sextama 框(Sextama Box) § -35序列,RNA聚合酶的松弛(初始)结合位点, § RNA聚合酶依靠其σ亚基识别该位点 —识别位点(R位点) § 大多数启动子中共有序列为 T82T84G78A65C54A45 § 重要性:很大程度上决定了启动子的强度 (RNApol 的σ因子) §位置在不同启动子中略有变动
保守T (2) Pribonow 框(Pribonow Box) § -10序列,RNA聚合酶的牢固结合位点 结合位点(B 位点) § 一致序列:T80A95T45A60A50T96 (TATPUAT) 因此又称TATA Box § 位置范围 -4 到-13
(3) 起始位点(initiation site):+1位点 RNA聚合酶的转录起始位点 起始NTP多为ATP或GTP
起始过程 : a. 全酶与启动子结合的封闭型启动子复合物的形成 (R位点被σ因子发现并结合 )
b、 开放型启动子复合物的形成 § RNApol的一个适合位点到达-10序列区域,诱导富 含A·T的Pribnow 框的“熔解”, 形成12-17bp的泡状 物,同时酶分子向-10序列转移并与之牢固结合 § 开放型启动子复合物使RNApol聚合酶定向 § 两种复合物均为二元复合物(全酶和DNA )
G C T A ---6-9 bp--- TTGACA -----16-18 bp------- TATAAT -35 sequence -10 sequence +1 c. 在开放型的启动子复合物中,RNApol的I位点和E位点的 核苷酸前体间形成第一个磷酸二酯键(β亚基) 三元复合物形成 +1位多为CAT模式,位于离开保守T 6~9 个核苷酸处 (4) σ因子解离→核心酶与DNA的亲和力下降 起始过程结束→核心酶移动进入延伸过程
转录的起始过程 σ 核心酶 12~17bp
3、 CAP-cAMP 结合位点 (也称CAP位点) 转录调控中,I → 阻遏蛋白 → 负调控 lac操纵子模型
许多基因的表达还存在正调控机制 例如: E.coli 培养基中乳糖和葡萄糖共存时 → 只有葡萄糖被 利用 原因:CAP位点的正调控 • 葡萄糖缺少时 ,腺苷酸环化酶将ATP → cAMP, cAMP与CAP位点结合,此时启动子的进入位点才能 与RNApol结合 • 葡萄糖存在时,cAMP不能形成,没有CAP-cAMP 与CAP位点结合,启动子的RNApol进入位点不能结 合RNApol
因此,一个操纵元中有两道控制开关,只有同时打开,结构基因才 能进行转录。 (对 lac操纵元而言,只有没有葡萄糖,有乳糖时才能进行录) (1)CAP位点的组成 (-70 ~ -40) 位点Ⅰ:-70 ~ -50 包括一个IR 序列 强结合位点 位点Ⅱ:-50 ~ -40 弱结合位点 位点Ⅰ对位点Ⅱ具有协同效应(cooperativity)
(2) 位点Ⅱ结合CAP-cAMP复合物后,促使RNApol进入 Sextama Box,最终与-10序列结合起始转录 原因:CAP-cAMP复合物与位点Ⅱ结合 → Sextama Box 富含G·C区域(G·C岛区)稳定性降低 → Pribnow Box 的溶解温度降低 →促进开放型启 动子复合物的形成 →促进转录
4、RNApol 在DNA上结合位点的鉴定 DNase 法----------40bp 而足迹法(footprinting)结果相对准确 足迹法原理: • 限制酶切割结合有RNApol的DNA →大分子DNA • 末端标记该DNA(Klenow片段标记3’,碱性磷酸酯 酶标记5’) • 用内切酶降解DNA(控制反应条件) • 凝胶电泳分离,放射自显影观察
限制酶切 分离 大片段DNA
末端标记大分子DNA 重新结合RNApol 作对照 直接用DNase 进行降解 用微量DNase降解 电泳 电泳
• 用足迹法鉴定出来的 RNApol 与DNA的结合区域为 +20 ~ -50(-40),大约 60 ~ 70 bp • 实验中还发现…………….. 5、 启动子各位点与转录效率的关系 (1)-35 序列与-10 序列与转录效率的关系 标准启动子 -35 TTGACA -10 TATAAT
则不同的启动子 a. 与标准启动子序列同源性越高 →启动强度越大 b. 与标准启动子同源性越低 →启动强度越小 c. 与标准启动子差异很大时 →由另一种σ因子启动 原因: • -35序列通过被 σ因子识别的容易 → 决定启动子强度 • -10序列影响开放型启动子复合物形成的速度 → 决定启动子强度
• 启动子上升突变、启动子下降突变 (2) -35序列与-10序列的间隔区与转录效率的关系 ◆ 碱基序列并不重要 ◆ 间距非常重要,17bp的间距转录效率最高 ◆ 间距上的突变种类: 间距趋向于17bp → 上升突变 间距远离17bp → 下降突变
E.Coli各σ识别的启动子的序列 δ24 δ54 δ28
第三节 真核生物RNA转录的启动 RNA Transcription promotion in Eukaryots
一、真核生物的RNApol 1、 三种RNApol: 根据对α - 鹅膏蕈碱的敏感性不同而分类 RNApolⅠ 最不敏感 (动、植、昆) RNApol Ⅱ 最敏感 RNApol Ⅲ 不同种类(物种)的敏感性不同 2、 位置和转录产物 RNApolⅠ核仁 活性所占比例最大 转录rRNA(5.8S、18S、 28S)
RNApolⅡ核质 主要负责 hnRNA、snRNA的转录 hnRNA(mRNA 前体,核不均一RNA heterogeneous nuclear RNA) snRNA(核内小分子 RNA small nulear RNA) RNApol Ⅲ核质 负责 tRNA、5S rRNA、Alu序列和 部分 snRNA •目前在线粒体和叶绿体内发现少数 RNApol(活性较低)
3、亚基组成: 分子量500KDa,含两个大亚基和 7~12 个(4-8)小亚基 RNApolⅡ: • 大亚基中有 C 末端结构域 (carboxy terminal domain CTD) • CTD中含一保守氨基酸序列的多个重复 Tyr-Ser-Pro-Thr-Ser-Pro-Ser C 端重复七肽 • 不同生物中重复数目不一样(酶活性)
• CTD中的 Ser和 Thr可被高度磷酸化 磷酸化的 RNApol Ⅱ被称为ⅡA 非磷酸化的 RNApol Ⅱ称为ⅡB • CTD参与转录 → ⅡB → ⅡA → 使 RNApol易于离开 启动子进入延伸过程(10倍) 二、 真核生物的启动子 三种 RNApol → 三种转录方式 三种启动子 → 三类基因,Ⅰ类 Ⅱ类 Ⅲ类