Download
1 / 45
460 likes | 508 Vues
Explore the principles of bioinorganic chemistry focusing on dioxygen activators such as P-450 and MMO with examples of atom and group transfer chemistry. Discover the key concepts related to substrate binding, redox changes, metal coordination, and electron transfer strategies.

E N D
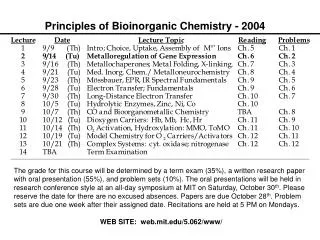
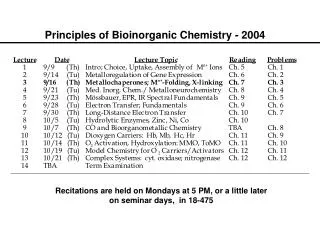
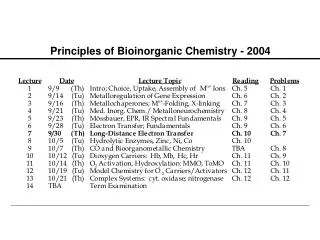
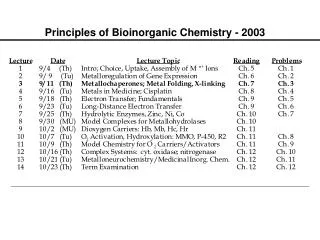
Principles of Bioinorganic Chemistry - 2004 Note: The course seminar presentations will be held on Sunday, October 31, 2004 beginning at 8:30 A.M. in the Bush Room. Please remember that daylight savings time ends that day.
Dioxygen Activators: P-450 and MMO Examples of Atom- and Group-Transfer Chemistry PRINCIPLES: • Both substrate binding and redox changes occur • Coupled proton-electron transfer steps set the redox potentials • Closely positioned redox/acid-base units work in concert • Interactions with substrates/other proteins gate electron transfer • Two-electron transfer strategies include 2 metals, M-porphyrins • Metal centers used to create or destroy radical species • Changes in metal coordination spheres can facilitate allostery • Bioinorganic chemistry of dioxygen paramount example ILLUSTRATIONS: • O2 Binding and Transport: hemoglobin (Hb), myoglobin (Mb), hemocyanin (Hc), and hemerythrin (Hr) • O2 Activation: cytochrome P-450, tyrosinase, methane monooxygenase; dioxygenases
Principles Illustrated by these Cases Substrate binding and redox changes occur: In all three cases, O2 binding is accompanied by electron transfer from one or two metal ions to dioxygen. Coupled proton-electron transfer steps set the potentials: In oxyHr a proton transfers from the bridging hydroxide to the peroxo ligand; this step appears to block further conversion to high-valent iron oxidase center(s). Metal center used to create or destroy radical species: Occurs in ribonucleotide reductase R2 protein. Changes in metal coordination sphere facilitate allostery: Explains the cooperativity of O2 binding in Hb.
The Cytochrome P-450 Reaction Cycle When an axial site is available on the iron porphyrin, dioxygen can bind and/or be activated there. With proton-mediated reductive activation of the O2 molecule, a peroxo intermediate forms that converts to an FeIV=O species, the ferryl ion. The ferryl can oxidize hydrocarbons to alcohols, epoxidize olefins, oxidize amines to amine oxides and do related chemistry. P-450’s are liver enzymes necessary for metabolism and used to convert pro-drugs and pro-carcinogens to their active forms.
The Mineral Springs in Bath, England, Source of Methylococcus capsulatus (Bath) The Restitutive Contents of the WATER’s Concoctive Power: Solution of gaffes, chaos of Salts and mineral effluvia of subterranean expiration. It cleanses the body from all blotches, scurvical itchings and BREAKING OUTS WHATSOEVER!
Methanotrophs are Used in Bioremediation Prince William Sound, Alaska: After the Exxon Valdez oil spill, fertilizers were spread on the beaches and natural methanotrophs restored their pristine beauty. Plants recruit oil-detoxifying microbes, as discovered by scientists analyzing the recovery of the environment in the Persian Gulf region following the 1991 Gulf War. " In the root zone was a rich reservoir of well-known oil eating microbes... one family of which (Arthrobacter) accounted for fully 95 percent..." Science News, 148, 84 (August 5, 1995)
Evolutionary Relationships Between Multicomponent Monooxygenases Common Ancestor Phenol Hydroxylases Amo Alkene Monooxygenase Dimethyl Sulfde Hydroxylase Phenol Hydroxylase Soluble Methane Monooxygenases Four Component Alkene/ Aromatic Monooxygenases Methane Monooxygenase Butane Monooxygenase Toluene Monooxygenases Phenol Hydroxylases Isoprene Monooxygenases
Properties of Hydrocarbon Monooxygenases Containing Carboxylate-Bridged Diiron Centers Leahy, Batchelor, Morcomb, FEMS Microbiol. Rev.2003, 770, 1-31.
Properties of Methanotrophs pMMO, Cu sMMO, Fe • 5-50 Tg CH4/year consumed by soil methanotrophs (1-10% of • atmospheric CH4), converting this greenhouse gas to biomass. • 104 kcal/mol BDE for methane makes it a challenge to activate. • Controlled oxidation to methanol at moderate temperatures in neutral aqueous solution is a remarkable chemical feat. • 500 billion barrels crude oil equivalent in recoverable but remote natural gas deposits might be made available.
Carboxylate-Bridged Diiron Proteins Global Research Goals • What tunes the properties of the diiron centers? • What are the electron transport pathways? • What factors control dioxygen reactivity? • How is substrate specificity achieved? Objectives for the sMMO and Model Studies • Determine structures of all components and complexes • Understand hydroxylation and epoxidation reactions • Synthesize and characterize structural/spectroscopic models • Achieve selective oxidation and catalysis
Soluble Methane Monooxygenase (sMMO) Component Structures & Reaction Cycle
sMMO is a Multicomponent Enzyme • Reductase • MMOR • Uses FAD and [2Fe-2S] for electron transfer from NADH to MMOH • Hydroxylase • MMOH • a2 b2 g2 • Dinuclear iron active site in each a subunit • Hydroxylation chemistry • Regulatory Protein • MMOB • required for full activity
MMOH Dinuclear Iron Active Site Glu209 Glu243 Glu114 His246 His147 Glu144 Hox, (FeIII)2 Hred, (FeII)2 Both Hox and Hred are charge neutral; X-ray structures by Rosenzweig, Whittington, et al., 1993-present
The Catalytic Cycle of sMMO H2O NAD+ O2 NADH MMOR B MMOHred MMOHsuperoxo MMOHox B B H+ MMOHperoxo Mössbauer (d 0.66 mm s-1) UV-vis, 725, 410 nm MMOHQ Mössbauer (d 0.17 mm s-1) UV-vis, 420 nm; EXAFS CH3OH CH4 RH RH(O) B
B Reactions of CH3X Substrates with Q H+ MMOHQ Mössbauer (d 0.17 mm s-1) UV-vis, 420 nm; EXAFS CH3OH CH4
Reaction of Q with Methane by Double-Mixing SF kH/kD = 26 CH4 T = 20 ºC [H]red = 16.8 µM [CH4] = 0.50 mM kobs = 14.1(1) s-1 CD4
Mechanism for Methanol Formation E = 0.0 kcal/mol Q Methane Gherman, Dunietz, Whittington, Lippard & Friesner, J. Am. Chem. Soc. 2001, 123, 3836. Baik, Gherman, Friesner & Lippard, J. Am. Chem. Soc., 2002, 124, 14608.
First Electron Transfer for Methanol Formation 17.9 kcal/mol • First electron transfer occurs here and determines the barrier height; one Fe reduced to Fe(III) as O–H bond forms.
Mechanism for Methanol Formation This transition state is 1.3 kcal/mol uphill from the bound radical intermediate, affording a rate constant in accord with most radical clock substrate probe studies.
Electronic Details of Second Electron Transfer Acceptor Orbital Donor Orbital Bound Methyl Radical (b-Spin) Fe1 d-(x2-y2) (b-Spin-LUMO) ‘Mediator’ Orbital Oxo p(z) (doubly occupied) Baik, Gherman, Friesner & Lippard, J. Am. Chem. Soc.2002, 124, 14608.
Electronic Details of Second Electron Transfer •H–O rotation promotes intramolecular b-electron transfer from the oxo lone pair orbital to the metal-based LUMO. •The remaining radicaloid a-electron on the bridging oxo group has the correct spin to recombine with the b-electron on the substrate to form a s-bond.
Overall Energetics and Methanol Release 1.3 -69.7 E in kcal/mol
Reactions of Q with Substrates Reveal Complexities C2H6 CH4 C2D6 CD4 Puzzles: This result indicates that, for ethane, the rate-determining step in not C–H bond activation. Yet kobs is the same! Answers: For CH4, H atom abstraction is rate determining; for C2H6, binding is rate determining. The bond in C2H6 is weaker, lowering of the C–H bond activation energy by ~5.6 kcal/mol, from both experiment and theory.
kobs vs Nitromethane Concentration for Q Decay Solid circles, CH3NO2 Open circles CD3NO2 pH = 7, 20 C ; KIE, 8.1 Direct evidence for bound substrate in a Q reaction is facilitated by the high solubility of nitromethane. Ambundo, E. A.; Friesner, R. A.; Lippard, S. J. J. Am. Chem. Soc.2002, 124, 8770-8771.
Single Turnover of Qwith Nitromethane-d3 at 25°C by Stopped-Flow Infrared Spectroscopy Loss of nitromethane-d3 monitored by stopped-flow IR spectroscopy at 1548 cm-1; kobs 0.39 s-1 Loss of Q monitored by stopped-flow spectrophotometry at 420 nm; kobs 0.39 s-1 First direct monitoring of the hydroxylation of a methane-derived substrate in the sMMOH reaction pathway Muthusamy, M.; Ambundo, E. A.; George, S. J.; Lippard, S. J.; and Thorneley, R. N. F. J. Am. Chem. Soc.2003, 125, 11150-11151.
KIE for Reactions of Q with CH3X Substratesa CLASS I SUBSTRATES H atom abstraction rate-determining: CH4, CH3CN, CH3NO2 CLASS II SUBSTRATES Binding rate-determining: C2H6, CH3OH Ambundo, E. A.; Friesner, R. A.; Lippard, S. J. J. Am. Chem. Soc.2002, 124, 8770-8771.
Gherman, B. F., Lippard, S. J., Friesner, R. A., submitted, 2004
RH RH(O) Reactions of Substrates with Hperoxo MMOHperoxo Mössbauer (d 0.66 mm s-1) UV-vis, 725, 410 nm
Preliminary Evidence for Hperoxo Reacting with Substrates filled circles - propylene open circles – methane Hperoxo appears to react with propylene Low solubility of substrates limits experiment Could propylene accelerate the conversion of Hperoxo to Q? Valentine, A. M.; Stahl, S. S.; Lippard, S. J. J. Am. Chem. Soc.1999, 121, 3876-3887.
l 705 or 720 nm l 420 nm Conditions: T = 20 ºC, [H]red = 51.5 mM, [B] = 103 mM Ether concentration in excess and variable, 3 - 70 mM Hperoxo and Q Reactions with Ethyl Vinyl Ether The product of propylene reaction with Hperoxo is propylene oxide
Results for Hperoxo and Q with Ethyl Vinyl Ether Second Order Rate Constant k = 1500(100) M-1s-1 Second Order Rate Constant k = 223(10) M-1s-1 Rate constant for Hperoxo is significantly greater than for Q. Diethyl ether reacts with Q reacts but Hperoxo does not. Conclusion: Hperoxodoes react with substrates; others are under investigation. Beauvais, Bautista, and Lippard (2004), in preparation
Mechanisms for Epoxidation of Ethyl Vinyl Ether By Hperoxo: By Q: Hperoxo may react by 2-electron transfer and Q by single-electron transfer. In addition, there might be distinct binding and activation steps.
O2 B MMOHred MMOHsuperoxo MMOHox B B H+ MMOHperoxo RH CH3OH RH(O) CH4 B Substrate Access in MMOH and ToMOH
NADH FAD e- NAD+ S Fe Fe S Toluene /o-Xylene Monooxygenase from Pseudomonas stutzeri OX1 ToMOR Substrates: toluene, xylenes, benzene, napthalene, phenol, cresols, ethylbenzene, styrene, halogenated ethylenes ToMOB Fe2S2 e- Rieske regulation e- + O2 Fe Fe Fe Fe + H2O ToMOH ToMO has relaxed substrate specificity All components are expressed recombinantly in E. coli! Carfaro, V., et. al. Eur. J. Biochem.2002, 296, 5689-5699.
Crystallization of ToMOH 0.7 mm 2Fo-Fc map from SAD Phasing Crystal Facts: Space group P3121 Unit cell (Å) 182 x 182 x 67 Conditions: ± ToMOB ± NaN3 (antimicrobial) M. H. Sazinsky, J. Bard, A. Di Donato, & S. J. Lippard, J. Biol. Chem. 2004, 279, 30600-30610.
Crystal Structures of ToMOH and MMOH MMOH ToMOH g-subunits The positions of the subunit differs for the two hydroxylases. The folds of the and subunits are conserved; those of are not. The canyon region in ToMOH is larger with a different aperture. Fewer protomer contacts occur for ToMOH.
Active Site Structures of ToMOHox and MMOHox Fe-Fe distance 3.0Å Fe-Fe distance 3.0 Å ToMOHox + thioglycolate MMOHox + acetate ToMOH has OH- and RCO2- bridges similar to those of MMOH and T4MOH. Pikus, J. D. et. al. Biochemistry. (1996), 35, 9106-9119. Cadieux, E., et al. Biochemistry. (2002), 41, 10680-10691.
ToMOH has a Surface Accessible Channel Openings Opening 1 Fork Fe Opening 2 • Channel is 35-40 Å long, 6-10 Å wide • Big enough for substrates and products • Diiron center has direct access to surface
Universal Pathways for Small Molecule Access CH2Br2 in MMOH cavities 2 and 3 4-Bromophenol in the ToMOH channel • ToMOH and MMOH use the same pathways for substrate entrance and product egress. • The channels and cavities may be a universal feature of all BMMs.
Substrate Access at the Diiron Center Helix F F205 I239 Helix F F196 Helix C I110 L110 Helix C F176 F188 Helix E Helix B Helix B Helix E MMOH TomoH The ToMOH Fe2 center is more accessible to solvent. “The most prominent structural differences …is in an altered side chain conformation for Leu 110 at the active site cavity. We suggest that this residue serves as one component of a hydrophobic gate controlling access of substrates to and products from the active site.” -Rosenzweig, et al., Proteins1997 29, 141-152
H2O NAD+ NADH MMOR B MMOHred MMOHox B Electron/H+ Transfer in MMOH and ToMOH
Helix E N214 N214 Helix E E240 E240 T213 E243 E243 T213 BrEtOH Helix F -OH Helix F Fe1 E114 Fe1 Helix B H147 E114 Helix B H147 Helix C Helix C Testing the Role of an Absolutely Conserved Asparagine Hred Hox + Bromoethanol Asn 214 is 100% conserved among all multicomponent monoxygenases. Cloning potential offers a chance to test function of N202 in ToMO. Could the role be to facilitate electron transfer? Proton transfer?
WT Specific Activity (nmol/min/ mg) WT 19% N202A 25% N202A Steady-State Turnover of WT and N202AToMOH (E. R. Cadieux and S. J. Lippard, Unpublished results (2004)) Single turnover results for ToMOHred + O2 + ToMOD+ phenol reveal no difference between wild type and N202A mutant protein activities. Working hypothesis: E. T. or H+ pathway involves N202 in the ToMO system.