受体介导的胞吞作用 Receptor-Mediated Endocytosis
380 likes | 748 Vues
受体介导的胞吞作用 Receptor-Mediated Endocytosis. 生科院 02 级 1 班 郭政、闫佳洁. 前言. 受体介导的胞吞作用是一种特殊类型的胞吞作用,主要是用于摄取特殊的生物大分子。如不同的蛋白质,包括激素、生长因子、淋巴因子和一些营养物都是通过这种方式进入细胞。. 问题的提出. 胚胎发育起始于一个微小的的精子和一个更大的卵细胞的结合。卵细胞是由卵母细胞发展而来的,并且积累卵黄。而这些卵黄是在雌性个体中其他部位合成的。那么,这些高分子量的卵黄蛋白是怎么能够进入卵母细胞的呢?.

受体介导的胞吞作用 Receptor-Mediated Endocytosis
E N D
Presentation Transcript
受体介导的胞吞作用Receptor-Mediated Endocytosis 生科院02级1班 郭政、闫佳洁
前言 • 受体介导的胞吞作用是一种特殊类型的胞吞作用,主要是用于摄取特殊的生物大分子。如不同的蛋白质,包括激素、生长因子、淋巴因子和一些营养物都是通过这种方式进入细胞。
问题的提出 • 胚胎发育起始于一个微小的的精子和一个更大的卵细胞的结合。卵细胞是由卵母细胞发展而来的,并且积累卵黄。而这些卵黄是在雌性个体中其他部位合成的。那么,这些高分子量的卵黄蛋白是怎么能够进入卵母细胞的呢?
1964年,哈佛大学的Thomas Roth和Keith Porter报道了有关蚊子的卵黄进入卵母细胞的可能的机理。他们注意到在卵母细胞快速增长时期,在卵母细胞表面有许多压低的纹孔,数量呈戏剧性的增长。这些纹孔,是由原生质膜的凹陷构成的。在它们的内表面被一层粗糙的表层所覆盖。
在一项有远见的计划中,Roth和Porter假定卵黄蛋白是被特别的吸附在有被小窝(coated pits)的膜的外表面,并将作为有被小泡(coated vesicles)而吸入。这些有被小泡脱离粗糙的表层后,一个与另一个结合,使得它们变得更大。
第一次观察到有被小泡的结构是直到1969年,由大阪大学的Toku Kanaseki和Ken Kadota发现的。电子显微镜检测到天然的小泡部分与豚鼠大脑相隔离,显示出有被小泡被一个多边形的柳条编制物覆盖。(图1 )这些调查表明胞被能够起到控制在小泡的形成中原生质膜的包裹作用。 有被小泡表面的网格模型 高倍电子显微镜下的照片
在生化自然杂志中的第一份有关有被小泡的研究是由Barbara Pearse于1975年在英国剑桥大学的一次医学研究会议上发表的。Pearse揭露了一个过程。此过程是通过将猪大脑的小泡膜通过一个连续的蔗糖浓度梯度做离心,直到有被小泡达到纯化分离。有被小泡中的蛋白质通过SDS—聚丙烯酰胺凝胶电泳被溶解和分离。
这个结果表示被膜含有一个分子量大约在180000道尔顿的占优势的蛋白质种类。Pearse命名这个蛋白为网格蛋白(clathrin)。他通过对几种不同类型动物细胞的分离,都发现了有被小泡在形成中的与此相同的蛋白质(从分子量和多肽图考虑的)。这个结果表示被膜含有一个分子量大约在180000道尔顿的占优势的蛋白质种类。Pearse命名这个蛋白为网格蛋白(clathrin)。他通过对几种不同类型动物细胞的分离,都发现了有被小泡在形成中的与此相同的蛋白质(从分子量和多肽图考虑的)。
新的研究 • 在上述的研究之后,1973年,在达拉斯的Texxas Medical学校的实验室中,由Michael Brown和Joseph Goldstein又共同提出了一个新的研究方向。Brown和Goldstein开始对家族性血胆脂醇过多(familial hypercholesterolermia FH)这种遗传病产生了兴趣。
Brown和Goldstein开始了对FH的研究,他们分别取正常人和FH患者的皮肤所派生的成纤维细胞(fibroblast)进行培养,检测胆固醇(cholesterol)的代谢过程。他们发现在胆固醇合成中酶的控制率,HMG辅酶A还原酶可以在正常的成纤维细胞中被环境中的脂蛋白(lipoprotein)(例如LDL)抑制。(图2)Brown和Goldstein开始了对FH的研究,他们分别取正常人和FH患者的皮肤所派生的成纤维细胞(fibroblast)进行培养,检测胆固醇(cholesterol)的代谢过程。他们发现在胆固醇合成中酶的控制率,HMG辅酶A还原酶可以在正常的成纤维细胞中被环境中的脂蛋白(lipoprotein)(例如LDL)抑制。(图2)
测量正常成纤维细胞中的HMG CoA还原酶活性。 • 正方形表示在胚胎血清中有部分脂蛋白。 • 实心圆表示全部脂蛋白。 • 三角形表示非脂蛋白。 • 从图中明显看出脂蛋白可以降低酶的活性,而非脂蛋白对酶活性几乎没有影响。 血清百分数 蛋白质浓度
这样,在正常成纤维细胞生长的培养基质中加入LDL(low-density lipoprptein低密度脂蛋白)的添加物,会导致HMG辅酶A还原酶活性水平的降低,并且伴随成纤维细胞的胆固醇合成的减少。
在发生FH的成纤维细胞中测量HMG辅酶A还原酶水平,发现它们是正常成纤维细胞的40~60倍。另外,全部FH成纤维细胞中的酶活性不受在环境媒介中LDL出现的影响。(图3)在发生FH的成纤维细胞中测量HMG辅酶A还原酶水平,发现它们是正常成纤维细胞的40~60倍。另外,全部FH成纤维细胞中的酶活性不受在环境媒介中LDL出现的影响。(图3)
在第六天时,培养基被新鲜的培养基替代。其中含有5%人类缺脂蛋白的原生质。在第六天时,培养基被新鲜的培养基替代。其中含有5%人类缺脂蛋白的原生质。 • 在测量初期可以明显看出细胞有很低的酶活性,这是因为培养基中含有足够的含胆固醇的脂蛋白,所以细胞不需要再合成。 • 一旦培养基换成缺脂蛋白的基质,细胞就不能利用基质中的胆固醇,于是就增加了细胞中酶的数量以便自身合成。 • 而相反的,在FH中显示出无论存不存在脂蛋白,细胞中的酶都无反应。 实心圆表示培养于含有10%胚胎血清的成纤维细胞。 空心圆表示纯合FH患者的成纤维细胞。
环境中的脂蛋白是怎样影响人工培养细胞的细胞质中的酶活性呢?环境中的脂蛋白是怎样影响人工培养细胞的细胞质中的酶活性呢? 为了解释这个问题,Brown和Goldstein开始了对细胞和脂蛋白之间相互影响的研究。
他们向培养皿中加入了放射性标记(radioactively labeled)的LDL,培养皿中包含有从FH患者或正常人中获得的成纤维细胞。正常的成纤维细胞以高亲和力特性结合被标记的LDL分子,但是突变的细胞显示出实际上没有能力与这些脂蛋白分子结合。(图4)
圆形表示放射性[125I ]标记的LDL在37℃与正常的细胞结合。三角形表示与FH的细胞结合。 • 空心圆和三角为细胞在含有5mg/ml[125I]LDL缓冲液的250mg/ml非放射性LDL中生长。实心圆和三角则不含有非放射性LDL。 • 在没有添加非放射性LDL的细胞中很明显看出,正常细胞结合了一定数量的LDL,而FH患者细胞则不能结合。 • 而在添加了非放射性LDL的细胞中标记LDL大量减少。因为非标记脂蛋白与标记的LDL竞争结合。
这些结论表示出正常细胞有一个LDL的高特异性受体,这个受体在有缺陷的或者在FH病人的细胞中丢失。这些结论表示出正常细胞有一个LDL的高特异性受体,这个受体在有缺陷的或者在FH病人的细胞中丢失。
为了显像受体结合和内化作用(internalization)的过程,Brown和Goldstein与Richard Anderson合作,Richard Anderson利用电子显微镜对细胞的结构有所研究。
这个研究小组从正常的和带有LDL的FH中慢慢提取成纤维细胞,这个LDL被共价的连接于铁蛋白中。因为铁原子,铁蛋白分子可以分散成电子带,就可以在电子显微镜下被显像。这个研究小组从正常的和带有LDL的FH中慢慢提取成纤维细胞,这个LDL被共价的连接于铁蛋白中。因为铁原子,铁蛋白分子可以分散成电子带,就可以在电子显微镜下被显像。 • 当在4℃时,正常的成纤维细胞和LDL—铁蛋白慢慢形成。在这个温度下配体可以与细胞表面结合,但不能被内化吸收,LDL—铁蛋白微粒被发现结合于细胞表面。
最近的检测解释LDL微粒不是随便的分散在细胞表面,而是在原生质膜上形成一个固定的小的片段(0.5μm)。这个膜被一个“绒毛的”物质切割和覆盖。(图5)最近的检测解释LDL微粒不是随便的分散在细胞表面,而是在原生质膜上形成一个固定的小的片段(0.5μm)。这个膜被一个“绒毛的”物质切割和覆盖。(图5) 电子显微镜下观察的人成纤维细胞中结合了LDL的有被小泡。LDL在结合了高密度电子铁蛋白微粒中很明显。
在这些膜上的片段是有被小窝,最初被Roth和Porter描述,并且在各种细胞类型中被发现。尽管来自FH病人的的细胞的表面被发现有相似数量的有被小窝,但没有LDL—铁蛋白结合在这些突变细胞上。这结果支持了突变的FH等位基因翻译成受体,而这个受体并没有能力结合LDL。后来用电子显微镜对LDL—铁蛋白的内化作用进行研究,显示出胞吞作用的路径是这些脂蛋白的内化。在这些膜上的片段是有被小窝,最初被Roth和Porter描述,并且在各种细胞类型中被发现。尽管来自FH病人的的细胞的表面被发现有相似数量的有被小窝,但没有LDL—铁蛋白结合在这些突变细胞上。这结果支持了突变的FH等位基因翻译成受体,而这个受体并没有能力结合LDL。后来用电子显微镜对LDL—铁蛋白的内化作用进行研究,显示出胞吞作用的路径是这些脂蛋白的内化。
基于这些结论,这个小组假定受体结合LDL的内化作用是在有被小窝中LDL受体的某部位密切相关的。接着这个假定,如果一个LDL受体不能在有被小窝上固定的,它就不能传递它的结合配体给细胞的溶酶体(lysosome),并且这样将不能影响在细胞中胆固醇的合成。基于这些结论,这个小组假定受体结合LDL的内化作用是在有被小窝中LDL受体的某部位密切相关的。接着这个假定,如果一个LDL受体不能在有被小窝上固定的,它就不能传递它的结合配体给细胞的溶酶体(lysosome),并且这样将不能影响在细胞中胆固醇的合成。
这时,一个不同类型的突变的LDL受体被发现。LDL受体提供了这个新的缺陷(通过病人发生J.D突变所了解的)结合放射性标记的LDL的正常总数,受体结合脂蛋白不能被内化,所以就不能被传送至细胞质的溶酶体进行加工。这时,一个不同类型的突变的LDL受体被发现。LDL受体提供了这个新的缺陷(通过病人发生J.D突变所了解的)结合放射性标记的LDL的正常总数,受体结合脂蛋白不能被内化,所以就不能被传送至细胞质的溶酶体进行加工。
Anderson和他的合作者假定LDL受体是一个跨膜蛋白,正常的会固定的在有被小窝上,因为它的细胞质区域是与有被小窝的成份特别的结合。可能网格蛋白(但是后来有关联的,例如适应物的小亚基,正如下面讨论的)因为这个在细胞质区域的缺陷,J.D突变受体是不能在细胞的有被小泡固定。有这种突变的人表现出相同的表现型,因为病人的受体是不能结合到LDL上的。Anderson和他的合作者假定LDL受体是一个跨膜蛋白,正常的会固定的在有被小窝上,因为它的细胞质区域是与有被小窝的成份特别的结合。可能网格蛋白(但是后来有关联的,例如适应物的小亚基,正如下面讨论的)因为这个在细胞质区域的缺陷,J.D突变受体是不能在细胞的有被小泡固定。有这种突变的人表现出相同的表现型,因为病人的受体是不能结合到LDL上的。
后来的研究决定了正常的LDL受体是个有839个氨基酸的跨膜糖蛋白,并在蛋白的C末端有50个氨基酸从膜延伸至内部作为细胞质的一个区域。在对J.D突变受体的分析中显示出蛋白质包括了一个简单的氨基酸代谢:一个酪氨酸残基正常定位的807位置被半胱氨酸取代。这个简单的在氨基酸序列上的变化消除了蛋白质在有被小窝上变得集中的能力。后来的研究决定了正常的LDL受体是个有839个氨基酸的跨膜糖蛋白,并在蛋白的C末端有50个氨基酸从膜延伸至内部作为细胞质的一个区域。在对J.D突变受体的分析中显示出蛋白质包括了一个简单的氨基酸代谢:一个酪氨酸残基正常定位的807位置被半胱氨酸取代。这个简单的在氨基酸序列上的变化消除了蛋白质在有被小窝上变得集中的能力。
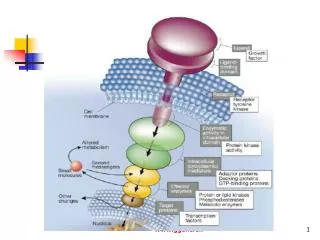
LDL与LDL受体结合,内吞入细胞使其获得脂类,主要是胆固醇,这种代谢过程称为LDL受体途径(LDL receptor pathway),该途径依赖于LDL受体介导的细胞膜吞饮作用完成 。
在接下来的几年中,他们的注意力转向了其他受体的细胞质尾部的氨基酸序列,这些受体在有被小窝中变得固定。那么,有没有一个共同的“内化作用信号”?有许多不同的受体比如胞吞作用的中间受体包括一个必要的酪氨酸残基,这些残基通常位于序列的YXXФ,这其中的Y是酪氨酸,而X可以是任意的氨基酸,而Ф是一个含有大的疏水链的氨基酸。研究表明受体的YXXФ序列与AP2结合素的一个多肽(μ亚基)结合。在接下来的几年中,他们的注意力转向了其他受体的细胞质尾部的氨基酸序列,这些受体在有被小窝中变得固定。那么,有没有一个共同的“内化作用信号”?有许多不同的受体比如胞吞作用的中间受体包括一个必要的酪氨酸残基,这些残基通常位于序列的YXXФ,这其中的Y是酪氨酸,而X可以是任意的氨基酸,而Ф是一个含有大的疏水链的氨基酸。研究表明受体的YXXФ序列与AP2结合素的一个多肽(μ亚基)结合。
X光射线显示出结合素和内化作用信号之间相互作用的特性。X光射线显示出结合素和内化作用信号之间相互作用的特性。 • 紫色表示膜受体上内化信号的X光下的结构。它结合在AP结合素的μ链上。 • 灰色表示μ亚基。 μ亚基几乎全部由β折叠片组成,有两个疏水的凹陷,其中一个与酪氨酸残基(Y)结合,另一个结合了内化作用信号的一个大的疏水侧链。L 为亮氨酸。
这种类型的相互作用就像是电线末端的两个分叉的插头与插座之间的对应关系。一旦内化作用信号的残基结合到结合素上,一种额外的吸引力被引起,通过在受体的中枢和μ亚基一段β片段之间结合氢的构造。这些不同分子间的联系的结果,结合素在有被小窝中被优先的胞吞作用捕捉。这种类型的相互作用就像是电线末端的两个分叉的插头与插座之间的对应关系。一旦内化作用信号的残基结合到结合素上,一种额外的吸引力被引起,通过在受体的中枢和μ亚基一段β片段之间结合氢的构造。这些不同分子间的联系的结果,结合素在有被小窝中被优先的胞吞作用捕捉。
我们再来回忆一下受体介导的LDL的胞吞作用的过程我们再来回忆一下受体介导的LDL的胞吞作用的过程
家族性血胆脂醇过多(familial hypercholesterolemia FH) 纯合体的个体含有致病基因(FH等位基因)会导致严重增加血清胆固醇浓度。(正常人为200mg/dl,而FH为800mg/dl),这种病常常导致阻断动脉(动脉粥样硬化),并且通常在患者20岁之前就死于心脏病。在当时,几乎没有人知道这种致病的生理原理。而且,目前尚无特效药物可以治疗。
LDL受体 • LDL受体是一种多机能蛋白,由839个氨基酸组成的36面体结构蛋白,分子量约115kD。由五种不同的区域构成,各区域有其独特的功能 。
LDL受体广泛分布于肝脏、动脉壁平滑肌细胞、肾上腺皮质细胞、血管内皮细胞、淋巴细胞、单核细胞、巨噬细胞,各组织或细胞的LDL受体活性差别很大。LDL受体广泛分布于肝脏、动脉壁平滑肌细胞、肾上腺皮质细胞、血管内皮细胞、淋巴细胞、单核细胞、巨噬细胞,各组织或细胞的LDL受体活性差别很大。 • LDL受体主要功能是通过摄取胆固醇进入细胞内,用于细胞增殖和固醇类激素及胆汁酸盐的合成等。
(1)血浆中LDL与细胞膜上有被区域的LDL受体结合。(1)血浆中LDL与细胞膜上有被区域的LDL受体结合。 • (2)使其出现有被小窝。 • (3)并从膜上分离形成有被小泡。 • (4)其上的网格蛋白解聚脱落,再结合到膜上。 • (5)其内的pH值降低,使受体与LDL解离。 • (6)LDL受体重新回到膜上进行下一次循环。 • (7)有被小泡与溶酶融合后,LDL经溶酶体酶作用,胆固醇水解。