Download

1 / 35
410 likes | 699 Vues
第七章 细胞信号传导( signal transduction ). 教师:李侠 学院:生命科学学院. 《 植物物生理学 》. 信号转导系统 一、概念 细胞偶联各种刺激信号与其引起的特定生理效应之间的一系列分子反应机制。. 细胞外. 质膜. 细胞内. G 蛋白连 接受体. 胞外信号. 受体. Ca/CaM;IP 3 /DAG 蛋白可逆磷酸化. 细胞反应. 二元组分系统. 信号输出. 信号转导网络. 信号输入. 跨膜信号转换. 二、过程. 第一节 信号与受体结合. 一、信号 (一)概念

E N D
第七章 细胞信号传导(signal transduction) 教师:李侠 学院:生命科学学院 《植物物生理学》 www.themegallery.com
信号转导系统 一、概念 细胞偶联各种刺激信号与其引起的特定生理效应之间的一系列分子反应机制。 www.themegallery.com
细胞外 质膜 细胞内 G蛋白连 接受体 胞外信号 受体 Ca/CaM;IP3/DAG 蛋白可逆磷酸化 细胞反应 二元组分系统 信号输出 信号转导网络 信号输入 跨膜信号转换 二、过程 www.themegallery.com
第一节 信号与受体结合 一、信号 (一)概念 对植物而言,环境变化就是刺激,就是信号。 (二)分类(根据性质) 物理信号:细胞感受到刺激后产生的能够传递信息作用的光、电和水力等信号。 化学信号(配体):细胞感受刺激后合成并传递到作用部位引起生理反应的化学物质(激素和病原因子)。 www.themegallery.com
受触及的含羞草小叶在1至2 秒钟向下弯,这是由于电波引发叶枕运动细胞中大量的K+和Ca2+转运,引起膨压改变的结果 www.themegallery.com
Albizia pulvini 背侧和腹侧的运动细胞之间的离子流调节了小叶的开放与闭合。 www.themegallery.com
植物激素是植物体主要的胞间化学信号。 如当植物根系受到水分亏缺胁迫时,根系细胞迅速合成脱落酸(ABA),ABA再通过木质部蒸腾流输送到地上部分,引起叶片生长受抑和气孔导度的下降。而且ABA的合成和输出量也随水分胁迫程度的加剧而显著增加。 www.themegallery.com
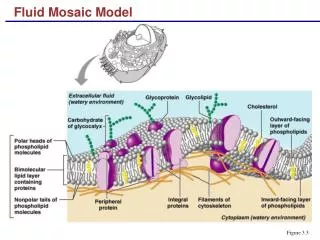
二、受体 (一)概念: 能够特异地识别并结合信号、在细胞内放大和传递信号的物质。 (二)特征: 特异性 高亲和力 可逆性 (三)类型 细胞表面受体 细胞内受体 www.themegallery.com
第二节 跨膜信号转换 一、概念 信号与细胞表面的受体结合之后,通过受体将信号传递进入细胞内的过程 二、两种方式 (一)G蛋白(GTP结合调节蛋白) 1.G蛋白的发现:吉尔曼(Gilman)、罗德贝尔(Rodbell) 获诺贝尔医学生理奖(1994) 2.细胞内的G蛋白一般分为两大类: 异源三聚体GTP结合蛋白:三种亚基(、、)构成 小G蛋白:一个亚基的单体 www.themegallery.com
1 2 G蛋白位于膜内侧,并与质膜紧密结合。 • 某种刺激信号与其膜上的特异受体结合后,激活的受体将信号传递给G蛋白, • G蛋白的α亚基与GTP结合而被活化。活化的α亚基与β和γ亚基复合体分离而呈游离状态, • 活化的α亚基继而触发效应器(如磷酸脂酶C) 把胞外信号转换成胞内信号。 • 而当α亚基所具有的GTP酶活性将与α亚基相结合的GTP水解为GDP后,α亚基恢复到去活化状态并与β和γ亚基相结合为复合体。 这样完成一次循环。 4 3 异源三体G蛋白的活动循环 www.themegallery.com
3.G蛋白作用机理G蛋白的信号偶联功能是靠GTP的结合或水解所产生的变构作用来完成的。当G蛋白与受体结合而被激活时,它就同时结合上GTP,继而触发效应器,把胞间信号转换成胞内信号;而当GTP水解为GDP后,G蛋白就回到原初构象,失去转换信号的功能。4.功能G蛋白参与细胞分裂、气孔运动、花粉管生长等生理反应。 小G蛋白参与细胞骨架的运动、细胞扩大、根毛发育以及细胞极性生长。 www.themegallery.com
(二)双元系统 1.受体分两个基本部分 组氨酸蛋白激酶(HPK) 反应调节蛋白(RR) 2.机理 3.例证:乙烯的受体就是双元系统 www.themegallery.com
初级信号(第一信使):胞外各种刺激信号; 次级信号(第二信使second messenger):由胞外刺激信号激活或抑制的、具有生理调节活性的细胞内因子。 植物中的的第二信使,主要是Ca2+、 cAMP、cGMP、DAG 、IP3 等。 第三节 细胞内信号转导形成网络 www.themegallery.com
一、Ca/CaM在信号转导中的作用 (一) Ca2+在细胞中的分布: 细胞质中Ca2+浓度约10-7~10-6mol·L-1,胞外Ca2+浓度约为10-4~10-3mol·L-1;胞壁是细胞最大的Ca2+库,其浓度可达1~5mol·L-1;细胞器的Ca2+浓度也是胞质的几百到上千倍,所以Ca2+在植物细胞中的分布极不平衡。 www.themegallery.com
胞外刺激信号可能直接或间接地调节这些钙离子的运输系统,引起胞内游离钙离子浓度变化以至影响细胞的生理生化活动。胞外刺激信号可能直接或间接地调节这些钙离子的运输系统,引起胞内游离钙离子浓度变化以至影响细胞的生理生化活动。 www.themegallery.com
几乎所有的胞外刺激信号(如光照、温度、重力、触摸等物理刺激和各种植物激素、病原菌诱导因子等化学物质)都可能引起胞内游离钙离子浓度的变化,而这种变化的时间、幅度、频率、区域化分布等不尽相同,所以有可能不同刺激信号的特异性可靠钙离子浓度变化的不同形式来体现。几乎所有的胞外刺激信号(如光照、温度、重力、触摸等物理刺激和各种植物激素、病原菌诱导因子等化学物质)都可能引起胞内游离钙离子浓度的变化,而这种变化的时间、幅度、频率、区域化分布等不尽相同,所以有可能不同刺激信号的特异性可靠钙离子浓度变化的不同形式来体现。 许多信号如蓝光、触摸能改变膜势和活化通道,使钙能进入,增加胞质中Ca+,这样引起K+和Cl+通道的打开,失去膨压,这种分别的转导途径通过 Ca+/钙调蛋白激酶和其他蛋白联系起来。蓝光和触摸也能通过其他路线改变细胞内的Ca+、 www.themegallery.com
Ca2+信号反应在气孔关闭过程中,与多个离子通道的活性及H+泵相协调Ca2+信号反应在气孔关闭过程中,与多个离子通道的活性及H+泵相协调 ①受体(R)感受ABA的作用导致了Ca2+的输入或Ca2+从内部储存中的释放, ②细胞质中的自由Ca2+浓度提高,促进了质膜上阴离子与K+Out通道的开放,并抑制了K+in通道的开放。 ③离开细胞的离子比进入细胞的多时,流出细胞水的量就会超过进入的水,如此,细胞就会失去膨胀作用,从而使得气孔关闭。 www.themegallery.com
(二)钙结合蛋白 类型:钙调素、钙依赖型蛋白激酶、钙调磷酸酶B相似蛋白 钙调素(calmodulin,CaM)是最重要的多功能Ca2+信号受体,由148个氨基酸组成的单链的小分子(分子量为17 000~19 000)酸性蛋白。 CaM分子有四个Ca2+结合位点。 机理:当外界信号刺激引起胞内Ca2+浓度上升到一定阈值后(一般≥10-6mol), Ca2+与CaM结合,引起CaM构象改变。活化的CaM又与靶酶结合,使靶酶活化而引起生理反应。 www.themegallery.com
目前已知有十多种酶受Ca2+-CaM的调控,如多种蛋白激酶、NAD激酶、H+-ATPase、Ca2+-ATP酶、 Ca2+通道等。 • 在以光敏色素为受体的光信号传导过程中Ca2+-CaM的信号系统也起着重要的调节作用。 • 生长素、光、摩擦等都可引起CaM基因活化,使CaM含量增加。 www.themegallery.com
二、IP3/DAG在信号转导中的作用 肌醇磷脂是一类由磷脂酸与肌醇结合的脂质化合物。主要以三种形式存在于植物质膜上:磷脂酰肌醇、磷脂酰肌醇-4-磷酸(PIP)、和磷脂酰肌醇-4,5-二磷酸(PIP2)。 www.themegallery.com
IP3作为信号分子,在植物中它的主要作用靶为液泡,IP3作用于液泡膜上的受体后,可影响液泡膜形成离子通道,使Ca2+从液泡这一植物细胞中重要的贮钙体中释放出来,引起胞内Ca2+浓度提高,从而启动胞内Ca2+信号系统来调节和控制一系列的生理反应。IP3作为信号分子,在植物中它的主要作用靶为液泡,IP3作用于液泡膜上的受体后,可影响液泡膜形成离子通道,使Ca2+从液泡这一植物细胞中重要的贮钙体中释放出来,引起胞内Ca2+浓度提高,从而启动胞内Ca2+信号系统来调节和控制一系列的生理反应。 www.themegallery.com
DAG的受体是蛋白激酶C(PKC),在一般情况下,质膜上不存在自由的DAG。在有DAG、Ca2+时,磷脂与PKC分子相结合,PKC激活,使某些酶类磷酸化,导致细胞反应;当胞外刺激信号消失后,DAG首先从复合物上解离下来,而使酶钝化,与DAG解离后的PKC可以继续存在于膜上或进入细胞质里备用。DAG的受体是蛋白激酶C(PKC),在一般情况下,质膜上不存在自由的DAG。在有DAG、Ca2+时,磷脂与PKC分子相结合,PKC激活,使某些酶类磷酸化,导致细胞反应;当胞外刺激信号消失后,DAG首先从复合物上解离下来,而使酶钝化,与DAG解离后的PKC可以继续存在于膜上或进入细胞质里备用。 大载二萜醇酯 细胞蛋白的磷酸化(作用) www.themegallery.com
双信号系统: 以肌醇磷脂代谢为基础的细胞信号系统,是在外信号被膜受体接受后,以G蛋白为中介,由质膜中的磷脂酶 C(PLC)水解PIP2,生成肌醇-1,4,5-三磷酸(IP3)和二酰甘油(DAG)两种信号分子。 IP3通过调节Ca2+浓度,而DAG则通过激活蛋白激酶C(PKC)传递信息。 www.themegallery.com
三、信号转导中的蛋白质可逆磷酸化 蛋白激酶PK NDP NTP 蛋白质 蛋白-PI H2O 蛋白磷酸酶PP PI www.themegallery.com
四、蛋白质的磷酸化和去磷酸化 植物体内许多功能蛋白转录后需经共价修饰才能发挥其生理功能,蛋白质磷酸化就是进行共价修饰的过程。 蛋白质磷酸化以及去磷酸化是分别由一组蛋白激酶(protein kinase)和蛋白磷酸酯酶(protein phosphatase)所催化的。 蛋白质可逆磷酸化反应式 NTP代表ATP或GTP,NDP代表ADP或GDP, www.themegallery.com
(一)蛋白激酶 蛋白激酶三类:丝氨酸/苏氨酸激酶、酪氨酸激酶和组氨酸激酶,它们分别将底物蛋白质丝氨酸/苏氨酸、酪氨酸和组氨酸残基磷酸化。有的蛋白激酶具有双重底物特异性,既可使丝氨酸或苏氨酸残基磷酸化,又可使酪氨酸磷酸化。 www.themegallery.com
Ca2+ 激酶活性 钙依赖型蛋白激酶 蛋白激酶中研究得较多的是最初从大豆中得到的依赖于钙离子的蛋白激酶(calciumdependent protein kinase,CDPK),CDPK有一与钙结合位点。 一般来说, CDPK在其氨基端有一个激酶活性域,在其羧基端有一个类似CaM的结构区域,在两者之间还有一个抑制域。当位于CDPK上类似CaM的结构区域上的钙离子结合位点与Ca2+结合后,抑制被解除,酶就被活化。 www.themegallery.com
MAPK:促分裂原活化蛋白激酶 MAPKK MAPKKK MAPKKKK www.themegallery.com
大豆、玉米、胡萝卜、拟南芥等植物中都存在这类蛋白激酶。大豆、玉米、胡萝卜、拟南芥等植物中都存在这类蛋白激酶。 • 现已知的可被CDPK磷酸化的作用靶(或底物分子)有细胞骨架成分、膜运输成分、离子通道、质膜上的质子ATP酶等。 www.themegallery.com
(二)蛋白磷酸酶 • 蛋白磷酸酯酶逆磷酸化作用,是终止信号或一种逆向调节。 • 现在植物中鉴定出几种不同的蛋白质磷酸酯酶。如豌豆、胡萝卜中的Type1蛋白磷酸酯酶、豌豆、胡萝卜、小麦中的Type2A和Type2C蛋白磷酸酯酶等。 • 蛋白质磷酸化的可逆性为细胞的信息提供了一种开关作用。 • 有研究表明,Type1蛋白磷酸酯酶可能与植物细胞的有丝分裂过程的调控有关。 www.themegallery.com
细胞内信号转导的双信号系统 IP3作为信号分子,在植物中它的主要作用靶为液泡,IP3作用于液泡膜上的受体后,可影响液泡膜形成离子通道,使Ca2+从液泡这一植物细胞中重要的贮钙体中释放出来,引起胞内Ca2+浓度提高,从而启动胞内Ca2+信号系统来调节和控制一系列的生理反应。 www.themegallery.com
环境刺激 信号传导的分子途径 胞间信号传递 第一信使 胞间信号 细胞外 膜上信号转换 受体 膜上信号转换系统 细胞膜 G蛋白 效应器 胞内信号转导 第二信使 细胞内 DAG IP3 Ca2+ cAMP 蛋白质可逆磷酸化 PKC PK Ca2+ CaM PKCa PKA Ca2+CaM CaM结合蛋白 酶蛋白磷酸化修饰 细胞反应 www.themegallery.com
Thank You ! www.themegallery.com