Download

1 / 33
450 likes | 1.39k Vues
Aminoácidos, péptidos y proteínas. Dr. Carlos antonio rius alonso Depto. De quimica organica Facultad de quimica Unam septiembre 200. Estructura de una proteína. Los aminoácidos están unidos mediante "enlaces amida" denominados enlaces peptídicos.

E N D
Aminoácidos, péptidos y proteínas Dr. Carlos antonio rius alonso Depto. De quimicaorganica Facultad de quimica Unam septiembre 200
Estructura de una proteína Los aminoácidos están unidos mediante "enlaces amida" denominados enlaces peptídicos. Las propiedades físicas y químicas de las proteínas se determinan a partir de los aminoácidos que las forman.
La configuración (S) de los aminoácidos. Casi todos los aminoácidos naturales tienen configuración (S), con estereoquímica parecida a la del L-(-)-gliceraldehído, por lo que se denominan L-aminoácidos. Excepto la glicina, todos los α aminoácidos son quirales. El centro quiral es el átomo de carbono asimétrico α.
Aminoácidos estándar. Hay veinte α-aminoácidos, denominados aminoácidos estándar, que prácticamente se encuentran en todas las proteínas.
Aminoácidos estándar. Los aminoácidos estándar difieren unos de o tros en la estructura de las cadenas laterales enlazadas a los átomos de carbono α.
Formación zwitterión. • A pesar de que generalmente los aminoácidos se escriben con un grupo carboxílico (-COOH) y un grupo amino (-NH2), su estructura real es iónica y depende del pH. • El grupo carboxílico pierde un protón, dando lugar a un ión carboxilato, y el grupo amino se protona y da lugar a un ión amonio. A esta estructura se le denomina ión dipolar o zwitterión.
Curva de valoración de la glicina El pH controla la carga de la glicina: catiónica por debajo de pH = 2.3; aniónica por encima de pH = 9.6 y zwitteriónica entre pH = 2.3 y 9.6. El pH isoeléctrico es 6.0. El punto isoeléctrico es el pH al que el aminoácido se encuentra en equilibrio entre las dos formas, como el zwitterión dipolar con una carga neta de cero.
Separación electroforética. Representación simplificada de la separación electroforética de la alanina, lisina y ácido aspártico a pH = 6. La lisina catiónica es atraída hacia el cátodo, el ácido aspártico aniónico es atraído hacia el ánodo y la alanina se encuentra en su punto isoeléctrico, por lo que no se mueve
Síntesis de Gabriel y malónica. Uno de los mejores métodos para sintetizar aminoácidos consiste en combinar la síntesis de Gabriel de aminas con la síntesis malónica de ácidos carboxílicos. El éster N-ftalimidomalónico se alquila de la misma forma que el éster malónico. La hidrólisis y la descarboxilación da lugar a un α-aminoácido racémico.
Síntesis de Strecker de alanina. Mediante la síntesis de Strecker se puede obtener una gran variedad de aminoácidos a partir de aldehídos apropiados. El mecanismo es similar al de la formación de una cianohidrina, excepto en que en la síntesis de Strecker el ión cianuro ataca a la imina en lugar de al aldehído.
Mecanismo de la síntesis de Strecker. Primero, el aldehído reacciona con amoniaco para formar una imina. La imina es un análogo nitrogenado del grupo carbonilo y es electrofílica cuando se protona. El ataque del ión cianuro a la imina protonada da lugar a α-aminonitrilo. En un paso separado, la hidrólisis del α-aminonitrilo da lugar a un α-aminoácido
Enzima acilasa. Una enzima acilasa (como la acilasa del riñón de cerdo o la carboxipeptidasa) sólo desacila al aminoácido natural L La mezcla resultante de D-aminoácido y de L-aminoácidos desacilados se separa fácilmente
Reacción de un aminoácido con ninhidrina. La ninhidrina es un reactivo común para visualizar las manchas o bandas de los aminoácidos que se han separado por cromatografía o electroforesis. La ninhidrina produce el mismo colorante púrpura, independientemente de la estructura del aminoácido original. La cadena lateral del aminoácido se pierde en forma de aldehído. La ninhidrina se puede utilizar en la detección de huellas dactilares dado que existen trazas de aminoácidos en las secreciones de la piel
Estabilización por resonancia. la estabilización por resonancia de una amida da lugar a su gran estabilidad, a la disminución de basicidad del átomo de nitrógeno y a la rotación restringida (carácter de doble enlace parcial) del enlace C-N. En un péptido, el enlace amida se denomina enlace peptídico. Tiene seis átomos en un plano: el C y el O del carbionilo, el N y su H, y los dos átomos de carbono asociados.
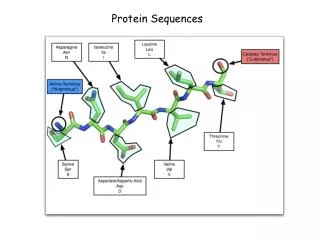
Hormona humana bradiquinina. Hormona humana bradiquinina es un nonapéptido con un grupo -NH3+ en el extremo N terminal y un -COO- libre en el extremo C terminal. Un péptido es un polímero de aminoácidos unidos por enlaces amido entre el grupo amino de cada aminoácido y el grupo carboxilo del aminoácido vecino.
Uniones disulfuro. La cistina, dímero de la cisteína, se obtiene cuando dos residuos de cisteína se oxidan y forman un puente disulfuro. La oxidación suave enlaza dos moléculas de un tiol para formar un puente disulfuro.
Oxitocina humana. La oxitocina es un nonapéptido con dos residuos de cisteína (en las posiciones 1 y 16) que unen parte de la molécula formando un gran anillo
Insulina bovina. La insulina está formada por dos cadenas peptídicas separadas, la cadena A contiene 21 residuos de aminoácidos y la cadena B que contiene 30.
Oxidación de una cistina a ácido cisteico La oxidación de una proteína mediante la ruptura de todas las uniones disulfuro por el ácido peroxifórmico, oxidando la cisteína a ácido cisteico. Los puentes de cisteína, cuando se reducen a la forma tiol, tienen tendencia a reoxidarse y volver a formar puentes disulfuro.
Determinación de la composición de los aminoácidos. En el analizador de aminoácidos, el hidrolizado pasa a través de una columna de intercambio iónico. La solución que emerge de la columna se trata con ninhidrina y su absorbancia se registra en función del tiempo. Cada aminoácido se identifica por el tiempo de retención necesario para pasar a través de la columna.
Composición de la bradiquinina humana Utilización de un analizador de aminoácidos para determinar la composición de la bradiquinina humana. Los picos de la bradiquinina para la Pro, Arg y Phe son más grandes que los de la mezcla equimolecular estándar, ya que la bradiquinina tiene tres residuos Pro, dos residuos Arg y dos residuos Phe
Secuenciación de la oxitocina bovina Los dos primeros pasos de la secuenciación de la oxitocina bovina se ilustran en esta figura. En cada degradación de Edman se rompe el aminoácido del N-terminal y se forma su derivado de feniltiohidantoína. El péptido acortado está disponible para el paso siguiente.
El método de Sanger El método de Sanger para la determinación N-terminal es una alternativa menos frecuente a la degradación de Edman. La degradación de Edman se suele preferir al método de Sanger
Análisis de los residuos C-terminales El aminoácido C-terminal se puede identificar utilizando el enzima carbixipeptidasa, que rompe el enlace peptídico del residuo C-terminal. Ocasionalmente, se hidroliza el péptido completo en sus aminoácidos individuales.
Unión del aminoácido del extremo C-terminal. De hecho, el polímero sirve como la parte alcohólica de un grupo protector éster para el extremo carboxílico del aminoácido del extremo C-terminal. Una vez que el aminoácido del extremo C-terminal se ha fijado al polímero, la cadena se forma a partir del grupo amino de este aminoácido.
Ruptura del péptido final. Cuando se completa la síntesis, el enlace éster con el polímero se rompe con HF anhidro Debido a que se trata de un enlace éster, se rompe con más facilidad que los enlaces amido del péptido
Acoplamiento del DCC. Cuando se trata una mezcla de una amina y un ácido con N,N'-dicilohexilcarbodiimida (DCC), la amina y el ácido reaccionan, y forman una amida. La molécula de agua que se pierde en la condensación transforma el DCC en N,N'-diciclohexilurea (DCU).
Derivado de acilo activado. El ión carboxilato se adiciona al carbono fuertemente electrofílico de la diimida, dando lugar a un derivado de acilo activado
Acoplamiento. El derivado activado reacciona rápidamente con la amina para dar lugar a la amida. En el paso final, la DCU sirve como un grupo saliente excelente.
Cada grupo carbonilo peptídico se une mediante un enlace de hidrógeno a un hidrógeno del grupo N-H de la siguiente vuelta de la hélice. Las cadenas laterales se simbolizan por esferas verdes en el modelo molecular de la figura
Reordenamiento de lámina plegada. Cada grupo carbonilo peptídico está unido mediante enlace de hidrógeno al hidrógeno del grupo N-H de una cadena peptídica adyacente. Este ordenamiento puede hacer que se alineen muchas moléculas de péptidos unas al lado de las otras, dando lugar a una lámina bidimensional.
Estructura terciaria de las proteínas globulares. En la estructura terciaria de una proteína globular típica se mezclan segmentos de hélice α con segmentos de enrollamiento al azar en los puntos donde se dobla la hélice.
Estructuras de una proteína La estructura primaria es la estructura enlazada covalentemente, incluyendo la secuencia de aminoácidos y los puentes disulfuro. La estructura secundaria incluye las áreas de hélices α, láminas plegadas o enrollamientos al azar. En la estructura terciaria se incluye la conformación total de la molécula. En la estructura cuaternaria se incluye la asociación de dos o más cadenas peptídicas de la proteína activa.