Peptide Elongation
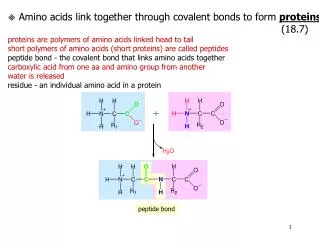
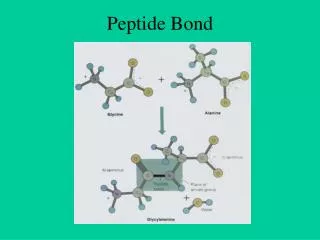
Peptide Elongation. Aminoacyl-tRNA-A-site biding. P. A. Peptide bond formation. This is catalyzed by a peptidyl transferase activity residing in the 23S rRNA. Evidence suggesting that 23S rRNA has peptidyl transferase activity:
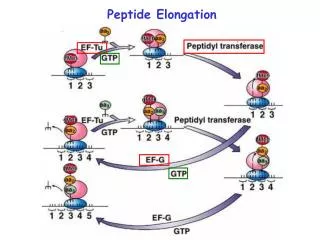

Peptide Elongation
E N D
Presentation Transcript
Peptide bond formation This is catalyzed by a peptidyl transferase activity residing in the 23S rRNA.
Evidence suggesting that 23S rRNA has peptidyl transferase activity: • Mutation in 23S rRNA, but not in any of the r-proteins, confer resistance to antibiotics that inhibit peptide bond formation. • Extraction of almost all the protein content of 50S subunit leaving <5% of r-proteins retains peptidyl transferase activity. However, treatments that damage RNA abolish the catalytic activity. • 23S rRNA prepared by in vitro transcription can catalyze the formation of a peptide bond, although with low efficiency.
Puromycin terminates protein synthesis by acting as an analogue of a tRNA charged with an aromatic amino acid.
NH2 NH NH Puro Puro Puro Inhibition of translation by puromycin Acid-insoluble Puromycin Peptidyl transferase Acid-soluble
Location of aa-tRNA and fMet-tRNAf can be determined by puromycin release assay fMet-tRNAf and Met-tRNAf can be released by puromycin aa-tRNA, like Met-tRNAm,can not be released by puromycin
Translocation This step requires EF-G plus GTP. GTP hydrolysis may cause a change in the structure of EF-G, which in turn forces a change in the ribosome structure. GTP hydrolysis is also needed to release EF-G. Models for translocation
Active centers in ribosome Peptidyl transferase EF-Tu/G-binding site
Change of 16S rRNA conformation during protein synthesis Triggered by joining with 50S subunit, binding of mRNA, binding of tRNA etc. Conformational change of ribosome during translocation is achieved mainly by alternative base-paring arrangement of rRNA.
Antibiotics that block prokaryotic protein synthesis: Streptomycin:inhibits peptide chain initiation and proofreading; increases misreading of pyrimidines. Tetracyclines:block aminoacyl-tRNA binding to the A site. Chloramphenicol:blocks peptidyl transferase. It is effective for bacterial and mitochondrial ribosomes. Erythromycin:inhibits translocation through the ribosome large subunit. The sites interacted with these antibiotics have been demonstrated to be located in the 16S rRNA (for streptomycin and tetracyclines) and 23S rRNA (for chloramphenicol) by primer extension assay used to define the region in the rRNA protected by the antibiotics.
Toxins that block protein synthesis: a-sarcin: an RNAase from Aspergillus that cleaves at a loop of eukaryotic large rRNA (corresponding the 23S rRNA of E. coli.) Ricin (produced by castor seeds): removes a base from eukaryotic large rRNA. Sites affected by these toxins are located in the same loop that is protected by elongation factors, EF-G and EF-Tu.
Comparison of EF-Tu and EF-G complexes Both EF-Tu and EF-G are ribosome-dependent GTPases. EF-Tu forms a ternary complex with tRNA and GTP, while EF-G forms a binary complex with GTP only. The structure of lower part of EF-G resembles the shape of tRNA in the ternary complex of EF-Tu. tRNA Aminoacyl-tRNA-EF-Tu-GDPNP EF-G-GDP
Molecular mimicry The ribosome-dependent GTPases, EF-Tu, EF-G, and RF3, are all structurally similar. Their binding sites in the ribosome may overlap, and this ensures that their binding with the 50S subunit is in an orderly manner.
Accuracy of elongationis achieved by: • Removing ternary complexes (aa-tRNA-EF-Tu) bearing the wrong aa-tRNA before GTP hydrolysis; • Eliminating the incorrect aa-tRNA before the wrong aa can be incorporated into the growing polypeptide chain (proofreading). • Both screens rely on the weakness of incorrect codon-anticodon base pairing to ensure dissociation will occur more rapidly before either GTP hydrolysis or peptide bond formation.
Accuracy of translationis achieved by: • Charging a tRNA only with the correct aa (a function of aminoacyl-tRNA synthetase, error rate: < 10-5); • Specificity of codon-anticodon recognition: proofreading by ribosome (error rate before proofreading: 10-1-10-2). • Factors affecting accuracy: • Geometry surrounding the A site affected by S12, S4, and S5. • Velocity of peptide bond formation. • Translation factors may also take part. • Overall accuracy of translation: ~ 5 X 10-4/codon
Genetic Code Ochre Opal Synonym codons: codons with same meanings. Amber The genetic code is universal, but exceptions exist. The set of tRNA responding to the various codons for each amino acid (codon usage) is distinctive for each organism.
Number of codons for each a.a. does not closely correlate with its frequency of use in proteins. E. coli
The degeneracy of the genetic code can be accommodated by isoaccepting species of tRNA that bind the same amino acid, or by wobble base pairing (non-Watson-Crick) between the codon and anticodon.
Wobble base pairs Standard pairing G-U wobble pairing
Preferential readings of modified bases for some codons may occur, e.g., uridine-5-oxyacetic acid and 5-methoxy- uridine recognize A and G more efficiently than U. U at the 1st position of the anticodon is usually converted to a modified form; A at that position is always converted to I. I pairs with C, U, and A, but not G. The first base of anticodon of Ile-tRNA, which recognizes AUA, AUU, and AUC, but not AUG, is I. The surrounding structure of anticodon also influences recognition of codons, because a change in a base in some other region of tRNA alters the ability of anticodon to recognize codons.
Termination of protein synthesis Termination (stop) codons UAA (ochre): most commonly used in bacteria. UGA (opal): causes more errors (1-3% are misread by Try-tRNA). UAG (amber) Release factors catalyze termination RF-1 recognizes UAA and UAG RF-2 recognizes UGA and UAA. RF-3, when binds GTP, helps RF-1 and RF-2 bind to and release from the ribosome. Cleavage of polypeptide from tRNA Use H2O instead of aminoacyl-tRNA as the acceptor of polypeptide. RRF (ribosome recycling factor), acts together with EF-G on 50S subunit to cause dissociation of 50S and 30S subunits.
Nonsense suppressor: stop codon suppression Wild-type Nonsense suppressor mutation Nonsense mutation
Nonsense suppressor tRNAs 1. Mutation in the anticodon (in E. coli) 2. Mutation outside the anticodon region For example, a G to A mutation at position 24 in the D stem of tRNATrp, which results in increased stability of the helix, allowing CCA to pair with UGA in an unusual wobble pairing of C with A, probably by altering the conformation of the anticodon loop.
Missense suppressor tRNA mutation The mutation can be suppressed by insertion either of the original aa or some other aa.
Effects of suppressor mutations In E. coli, amber suppressors tend to be relatively efficient (10-50%), but ochre suppressor are difficult to isolate and always much less efficient (< 10%). This difference may be because the ochre codon is used most frequently and suppression of this codon may be damaging to E. coli. Strong missense suppressor is not favored due to the damaging effects caused by a general substitution of aa. The effectiveness of a suppressor tRNA depends on the extent of its competition with the release factors or normal tRNA, which in turn is determined by the affinity between its anticodon and the target codon, its concentration in the cell, and other parameters. The extent of nonsense suppression by a given tRNA varies widely depending on the context of the codon. The base on the 3’ side of a codon have a strong effect.
Suppression of frameshift mutation Compensating base deletion or insertion; Suppressor mutations in tRNA tRNA recognizing a 4-base codon (e.g., change the anticodon of tRNAGly from CCC to CCCC). tRNA that blocks adjacent base by steric hindrance. Frameshifting as a normal event in natural translation Common features: A “slippery” sequence (aminoacyl-tRNA moves +1 or –1 base.) Ribosome is delayed at the frameshifting site by some ways to allow the aa-tRNA to rearrange its pairing. They include a scarce aminoacyl-tRNA recognizing the adjacent codon, a termination codon recognized slowly by its release factor, and a special conformation of RNA (“pseudonot”.)
Polysomes mRNAs are translated by multiple ribosomes in tandem.
Transcription and translation occur simultaneously in the bacteria Rates of transcription and translation are 40 nt/second and 15 aa/second, respectively. In one gene, there could be 5 initiations per minute and each mRNA may be translated by 30 ribosome.
Polycistronic mRNA Translation of polycistronic mRNA
Degradation of mRNA Half life of bacterial mRNA: ~2 min. mRNA degradation may be catalyzed by a complex that includes RNAase E (an endonuclease that makes the first cleavage for many mRNAs), polynucleotide phosphorylase (PNPase, a 3’-5’ exonuclease), and helicase. Secondary structure within mRNA may provide an obstacle to exonuclease, and this is unwound by the helicase. Some RNAs have a poly(A) tail (formed by the poly(A) polymerase) that acts as the binding site for the nucleases. The number of times an mRNA is translated is a function of the affinity of the SD region for ribosome and its stability.
Exceptional codons exist in mitochondria (fruit fly, mammalian, yeast, plant etc.) and the nuclear genome of ciliated protozoa or mycoplasma. Source Codon Usual meaning New meaning Fruit fly UGA Stop Tryptophan Mitochondria AUA Isoleucine Methionine Protozoa UAA Stop Glutamine Nuclei UAG Mycoplasma UGA Stop Tryptophan