Download

1 / 27
270 likes | 473 Vues
Lecture note series II. 2007 Pharmacology 4407B. Extracellular Ligand-Gated Receptors. Dr. Melanie Kelly Department of Pharmacology Dalhousie University. melanie.kelly@dal.ca. Extracellular Ligand-Gated Receptors. Belong to a family of homologous multi-pass transmembrane proteins

E N D
Lecture note series II. 2007 Pharmacology 4407B Extracellular Ligand-Gated Receptors Dr. Melanie Kelly Department of Pharmacology Dalhousie University. melanie.kelly@dal.ca
Extracellular Ligand-Gated Receptors • Belong to a family of homologous multi-pass transmembrane proteins • Activation of ELGR causes opening of the ion channel which forms a central pore through the receptor structure.
Properties of ELGR • They are activated in response to neurotransmitters and combine ion-selective functions with those for agonist binding. • They conduct ions through the impermeable cell membrane. • They select among different ions (specificity). • Changes in ion permeability following channel activation alter the excitability of cells. • Excitation of cells is associated with opening of cation-influx (depolarizing) channels, while inhibition of neuronal firing is generally associated with increased anion (Cl-) ion permeability and hyperpolarization.
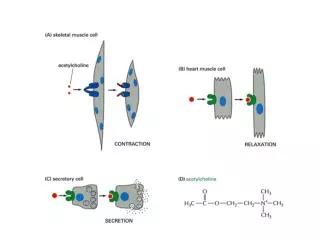
ELGR ACTIVATION Ligand-gated channels open in response to binding of a ligand. The pore of the ACh receptor channel is closed in the absence of ACh. Binding of the ligand (ACh) produces a conformational change that results in opening of the channel pore. Ions then move through the pore, resulting in single-channel currents.
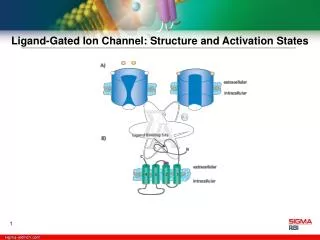
Organization of ELGR • The ligand-gated ion channel family consists of receptors composed of multiple subunits. • Each subunit has four transmembrane (TM) domains. Subunits exhibit sequence identities from 25 to 75% with a similar distribution of hydrophilic and hydrophobic domains. Topology of ELGR family A,B,C regions involved in agonist binding. N-terminus and C-terminus are extracellular Examples of ELGR receptors are: nAChR, 5HT3, Glycine receptors, GABAA receptor
Architecture of ELG Receptors Subunit topology (left) andsubunit organization (right) of two families of ELGR. Possible ligand binding sites are labelled with asterisks. The left hand diagram is of a single homologous subunit and the right hand diagram represents the subunit composition of the receptor
Functions of ELG Receptors • Fast excitatory transmission (nAChR) - Primary excitatory receptors in skeletal muscle and peripheral nervous system. • In central nervous system, nAChR are present in smaller numbers than glutamate receptors which mediate excitatory neurotransmission in both the central and peripheral nervous system. • GABA and Glycine are the main inhibitory neurotransmitters in the nervous system. Both ligands activate a receptor associated with chloride-selective channel.
Neuronal Nicotinic Acetylcholine Receptors (nAChR) • Sixteen genes encode the nAChR. 5 genes encode nAChR at the NMJ and the remaining 11 genes encode genes widely expressed in CNS and PNS.
Correlating Structure and Function of the nAChR • Structure of nAChR highly conserved, each receptor is pentamer with 5 subunits surrounding ionic pore. • Each subunit consist of aprox. 600 amino acids organized to span the membrane 4X. N- and C- termini lie in the synaptic cleft. The 2nd TM domain of each subunit forms the pore. • Neurotransmitter binding site is at interface of two adjacent subunits.
Pharmacology of nAChR • Drugs activating nAChR - drugs with structure similar to the endogenous neurotransmitter such that they can interact with binding site on the receptor e.g synthetic esters such as carbachol and nicotine. • Drugs blocking nAChR - bind to receptor on extracellular site and/or permeate the channel and bind in pore. Prevent ACh interacting with receptor e.g Tubocurare
Diseases Involving nACh Receptors • Myasthenia Gravis - autoimmune disease, prevalence of 7-9 cases/100,000 people. 2/3 patients are women, develops in early life with peak age of onset in 30’s. • Characterized by profound weakness of skeletal muscle which increases with exercise. Manifest initially in facial and eye muscles. Difficulty holding head up, chewing, swallowing, weakness in limbs. • Caused by autoantibodies directed at postsynaptic nACh receptors leading to loss of function. Mortality rare, treated with drugs that prolong the life-time of the neurotransmitter, ACh, in the synapse. Figure shows minature excitatory potentials recorded from normal (A) and Myasthenic human intercostal muscle (B). In the diseased muscle excitatory potentials due to nAChR activation are abnormally small and decreased in amplitude further with repetitive stimulation.
Neuronal nACHR and Epilepsy • Autosomal dominant frontal lobe epilepsy (ADFLE) is associated with a mutation in the CHRNA4 gene that encodes the 4 subunit of the high-affinity nACHR. • Mutation results in instability in neuronal cicuitary with excess excitation and abnormal synchrony. ADNFLE seizures arise mainly during stage II sleep in the frontal cortex and can progress to tonic-clonic seizures. • Penetrance of ADNFLE is incomplete with 70% of persons carrying the mutation displaying typical sleep pattern. • Mutant receptor has decreased channel open time, reduced single channel conductance and increased rate of desensitization. The reduction in nAChR function results in decreased activation of neurons releasing inhibitory neurotransmitter and results in enhanced excitability of postsynaptic neurons and lower seizure threshold.
Glutamate Receptors • L-glutamate, L-aspartate and other acidic amino acids act as excitatory neurotransmitters at synapses in the peripheral and central nervous system. • Glutamate primary excitatory neurotransmitter in CNS. • Two types of receptors: ionotropic and metabotropic. • Ionotropic are extracellular ligand-gated receptors (channels). • Metabotropic receptors do not have an integral ion channel and mediate their effects via G protein activation of second messenger cascade.
Ionotropic Excitatory Amino Acid Receptors • Different ionotropic glutamate receptors co-exist on many neurones classified according to their preferred agonist. Glutamate activates all channel subtypes. • N-methyl-D-aspartate receptors (NMDA)-slow kinetics, high Ca permeability. Important in generation of slow synaptic potentials and forms of activity-dependent synaptic plasticity • -amino-3-hydroxy-5-methyl-4-isoxazole propriate receptors (AMPA) -fast kinetics,Na+ permeability, usually low Ca permeability, rapid desensitization. Important in mediating rapid synaptic transmission (similarly for Kainate Receptors). • Kainate receptors - slow currents. Na+ permeability, Low Ca permeability.
Relationship between Genes Encoding different Glutamate Receptor Subunits • Line lengths are proportional to mean number of differences per residue along each branch.
Putative Topology of GluR Subunits • AMPA and Kainate subunits consist of three transmembrane domains (TMs 1,3,4), a large extracellular N-terminus and intracellular C-terminus. Pore is lined by hairpin loop (TM2) which enters from cytosolic side. • S1 and S2 domains are involved in agonist binding. • Flip/Flop domain is alternatively spliced module. • Q/R and R/G sites are subject to RNA editing (1st letter gives encoded by genomic DNA; 2nd letter indicates edited residue).
Functional Diversity • Heteromerization – Native AMPARs are predominantly heteromeric. Homomeric AMPARs are significantly more Ca2+ permeable than heteromeric channels. • mRNA editing – editing of mRNA results in substitution of arginine (R) residue for glutamine (Q). Designated Q/R site. When glutamine is present at Q/R site results in significant increases in Ca permeability of AMPARs. • Alternative Splicing – GluR1-GluR4 may exist in “flip” or “flop” version. Results from use of alternative exons to code for a region of 38 that are extracellular and preceed TM4. Flip/flop module affects desensitization rate of the channel. • Differential expression of flip/flop modules during development as well as between CNS and peripheral tissue.
NMDA Receptors • Binding of two glutamate (NMDA) and two glycine is required to activate NMDA channels. • Two major gene families encode NMDA receptor subunits: NR1 and NR2. • NR1 aprox 900 , 25% identity with AMPA and Kainate receptors, and is subject to alternative splicing with at 8 splice variants identified. • NR2 subunits (NR2A and NR2D) have longer C-terminal sequence and are larger (130 to 170 kDa). • Expression of NR1 and NR2 subunits is necessary for functional receptor and native NMDA receptors are thought to be composed of two NR1 and two NR2 subunits.
Activation of NMDA Receptors • Binding of glutamate or agonist (NMDA) to site on extracellular surface of receptor (thought to be NR2). • Glycine binding (to NR1) results in enhancement of the ability of glutamate or NMDA to open the channel through allosteric interaction. • Following binding of the agonist, depolarization results in removal of Mg2+ block and influx of Na+ and Ca2+ ions. • NMDARs more permeable to Ca2+ than other cations. • Influx of cations produces further depolarization.
Influx of Ca2+ through NMDARs can trigger long-lasting changes in synaptic efficacy.
Glutamate Receptors and Disease • Overactivation of GluR and excitotoxicity can occur due to pathological recurrent neuronal excitation i.e epilepsy, trauma, neurodegenerative diseases, ischemia or neurological toxicity - Demoic acid (kainate receptor agonist) found in mussels feeding on domoate-rich phytoplankton. • Seeds of chickling pea Lathyrus sativus - contain potent AMPA receptor agonist causes lathyrism, neurological disease with muscle ridgidity and spasm; • Flour made from seeds of cycad, Cycas Circulnalis -contains NMDA agonist causes Guam disease with symptoms of Parkinson’s and ALS.
GABA Type A Receptor • -aminobutyric acid (GABA) is the main inhibitory transmitter in the CNS. • Stimulation of inhibitory neurons in the CNS releases GABA onto adjacent neurons • Binding of GABA to receptors on the postsynaptic membrane causes a transient increase in permeability in Cl- via channel opening • The inhibitory actions of GABA are enhanced by the presence of barbiturates or benzodiazepines (allosteric modulators). • The GABAA Receptor is composed of and subunits and and/or subunits forming a functional receptor. The influx of Cl- in neurons causes hyperpolarization or inhibitory potentials which move the membrane potential of the neuron away from its firing threshold.
Inhibitory Neurotransmission • Antagonists of GABA receptors such as bicuculline and picrotoxin act as convulsants because they prevent GABA receptor activation. • Potentiators/agonists - enhance GABA receptor activation and depress the CNS, therefore are used as sedatives, anesthetics and anticonvulsants e.g barbiturates, benzodiazepines, volatile anesthetics.
Diseases/Syndromes Associated With GABA Channels • Angelman Syndrome - characterized by severe mental retardation, absence of speech, ‘puppet-like’ ataxic movements and seizures. • Incidence is 1 in 20,000 and majority of patients appear to be sporadic cases. • Disease in 70-80% of these patients associated with a deletion in region of maternal chromosome 15, such that the gene encoding GABAA subunit is deleted. Loss of GABAA subunit function contributes to neurological pathogenesis in this disease.