Download

1 / 45
520 likes | 945 Vues
Chapter 13 Membrane Channels and Pumps. Permeability of membrane is conferred by two classes of membrane proteins : PUMPs and CHANNELs. PUMP : use ATP energy or light absorption. = active transport .

E N D
Chapter 13 Membrane Channels and Pumps
Permeability of membrane is conferred by two classes of membrane proteins : PUMPs and CHANNELs. • PUMP : use ATP energy or light absorption. = active transport. • - CHANNEL : enable ions to flow through membranes in a thermodynamically downhill direction. = passive transport, or facilitated diffusion.
Pump : two types of ATP-driven pump, 1) P-type ATPase and 2)ATP-binding cassette(ABC) transporter. Pump can establishpersistent gradients of ions across membrane. • - Channel : can allow these ions to flow rapidly across membranes down gradient. The expression of transporters largely defines the metabolic activities of a given cell type • Each cell type express a specific set of transporters. • Transporters largely determine the ionic composition and the compounds inside cells cell’s characteristic reactions • Ex> GLUT(glucose transporter) have 5 homologs. • In case of GLUT3 of high affinity for glucose, expressed only on neurons and a few other cell types. When glucose is present at low concentration, first call on glucose.
13.1 The transport of molecules across a membrane may be active or passive - Two factors determine whether a molecule will cross a membrane 1) the permeability of the molecule in a lipid bilayer and 2) the availability of an energy source. Many molecules require protein transporters to cross membranes • Simple diffusion : molecule will pass through a membrane down their concentration gradient. Ex> lipophilic molecule, steroid hormones. • Passive transport : Charged ions pass through specific channels down their concentration gradient. Channels display substrate specificity. Ex> Na+ K+… • Active transport : Charged ion must be pumped against a concentration gradient(low conc. → high conc.) Energy is required.
Free energy stored in concentration gradients can be quantified • To abtain concentration gradient, requires an input of free energy because of a decrease in entropy. • Uncharged molecule ; • ΔG = RTln(c2/c1) = 2.303RTlog(c2/c1) • -Charged molecule ; • ΔG = RTln(c2/c1) + ZFΔV • = 2.303RTlog(c2/c1) + ZFΔV • R=gas constant • T=temperature in kelvins • Z=electrical charge • F=Faraday constant • ΔV= potential in volts across the membrane • A transport process must be active when ΔG is positive.
13.2 Two families of membrane proteins use ATP hydrolysis to pump ions and molecules across membranes • The ion gradients are generated by a specific transport system, an enzyme that is called the Na+-K+ pump or the Na+-K+ ATPase. • The hydrolysis of ATP by the pump provides the energy needed for the active transport of 3 Na+ out of the cell and 2 K+ into the cell, generating gradient (-50mV membrane potential maintenance). • 1/3 ATP consumed in resting animal • P type ATPase : ATPase form a key phosphorylated intermediate. • Phosphoryl group is linked to the side chain of a specific conserved aspartate residue in the ATPase. (SERCA, gastric H+-K+ ATPase)
P-type ATPases couple phosphorylation and conformational changes to pump calcium ions across membranes - Pump protein can exist in two principal conformational states, one with ion-binding sites open to one side of the membraneand the other with ion-binding sites open to the other side.
<Ca2+ ATPase = SERCA (example of P-type ATPase)> • In sarcoplasmic reticulum of muscle cell. • Constitutes 80% of the SR membrane protein. • Important role in muscle contraction. • Maintain Ca2+ concentration 0.1uM in cytoplasm while 1.5mM in SR • - Ca2+ bound SERCA. • Single 110kda polypeptide. • Transmembrane = 10 α helices. • 2 Ca2+ binding domains at TM. • 3 cytoplasmic domain. • N=ATP nucleotide binding • P=accept the phosphoryl group on a aspartate residue • A=actuator, linking changes in N & P domains to the transmembrane part
SERCA is structurally dynamic. • The SERCA without Ca2+ and with a phosphorylaspartate analog in the P domain. • The transmembrane part of the enzyme has rearranged and the well-organized Ca2+ binding site are disrupted. Ca2+ not bound Ca2+ bound
ATP bind. Rearragement of cytoplasmic domains. Phosphoryl group is transferred from ATP to Asp351. ADP release. Conformational change in membrane domain. Ca2+ release. Ca2+ bind. Enzyme everts back to the E1 conformation. Phosphoaspartate is hydrolyzed. Pi release.
Digitalis specifically inhibits the Na+-K+ pump by blocking its dephosphorylation • Na+-K+ pump a2b2 tetramer, a subunit homologous to SERCA • Certain steroid derived from plants are potent inhibitors. • Digitoxigenin : cardiotonic steroid. Inhibit the dephosphorylation of the E2-P form of the ATPase. • Digitalis : mixture of cardiotonic steroid from the dried leaf of the foxglove plant.
Digitalis increases the force of contraction of heart muscle. Inhibition of Na+-K+ pump → higher level of Na+ inside the cell → slower extrusion of Ca2+ by the Na+-Ca2+ exchanger → increase in the intracellular level of Ca2+ → enhance the ability of cardiac muscle to contraction. Congestive heart failure (울혈성심부전증) 치료약
P-type ATPase are evolutionarily conserved and play a wide range of roles Yeast : 16 proteins that belong to the P-type ATPase family (ca2+, Na+, Cu2+ ). ※ fippase : 5 members of this family. Transport of phospholipids with amino acid head group (ex, phosphatidylserine) membrane asymmetry Human : 70 proteins that belong to the P-type ATPase family.
Multidrug resistance highlights a family of membrane pumps with ATP-binding cassette domains • Multidrug resistance : the development of resistance to one drug also makes the cell less sensitive to a range of other compound. • Multidrug-resistance protein or P-glycoprotein : 170kda. ATP-dependent pump. • - MDR pump : 4 domains. • 2 membrane spanning + 2 ATP binding domain(=ATP-binding cassettes, ABCs). • Largest single family in the E.coli genome. Why? In Human? • Eukaryotes vs prokaryotes
Lipid transporter • Dimer of 62kDa chains. • N-term is membrane spanning • C-term is ATP-binding casette. • Two ABCs are in contact, but do not interact strongly in the absence of ATP. • P-loop NTPase family (GXXXXGK, 1st b strand and 1st helix) ABC transporter Structure from Vibrio
Substrate enters the central cavity. And conformational change in the ABCs.(increase affinity for ATP) Free of both ATP and substrate. ATP bind. Hydrolysis of ATP and release of ADP and Pi reset the transporter. Strong interaction between ABCs → conformational change in membrane spanning domain. Substrate out.
13.3 Lactose permease is an archetype of secondary transporters that use one concentration gradient to power the formation of another * cotransporter Couple the downhill flow of one species to the uphill flow of another in the opposite direction across the membrane. Use the flow of one species to drive the flow of a different species in the same direction across the membrane. Like channel, Transport a specific species in either direction governed only by concentrations of that species on either side of the membrane.
- Lactose permease of E. coli. Symporter. • Uses the H+ gradient across the membrane generated by the oxidation of fuel molecules to drive the uptake of lactose and other sugars against a concentration gradient. • Two helves, each of which contains 6 membrane spanning α helices. • Sugar lies in a pocket in the center of the protein.
A proton from outside the cell binds to a residue in the permease(Glu269). Permease binds lactose from outside the cell. Structure evert. Permease Evert. Permease release a proton to the inside of the cell. Permease release lactorse to the inside of the cell.
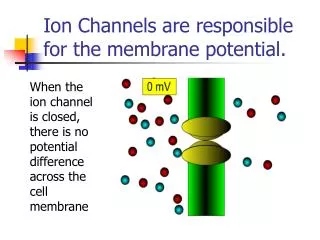
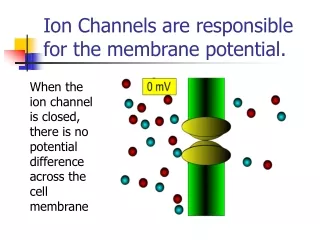
13.4 Specific channels can rapidly transport ions across membranes ※ Channel: Membrane protein . : Fast ion transport rate. : Highly sophisticated molecular machines. : Respond to chemical and physical changes and undergo precisely timed conformational changes.
Action potentials are mediated by transient changes in Na+ and K + permeability • Nerve impulse : electrical signal produced by the flow of ions across the plasma membrane of a neuron. • Resting state : High K+ concentration, low Na+ concentration inside of the cell(membrane potential : -60mV). • Depolarization : Na+ flow into the cell(Na+ channel open, membrane potential : +30mV). • Repolarization: K+ flow outward(K+ channel open, membrane potential : -60mV)
Patch-clamp conductance measurements reveal the activities of single channels To confirm the existence of the channel, ※Patch clamp technique : measure the ion conductance through a small patch of cell membrane. (by Erwin Neher and Bert Sakmann in 1976) Single ion channel undergoing transitions between closed and open states.
The activity of the channels in the entire plasma membrane can be monitored. Patch pipette diameter : 1μm A piece of plasma membrane with its cytoplasmic side now facing the medium is monitored. -The flow of ions through a single channel can be monitored with a time resolution of microseconds. -First views of single biomolecule activities
The structure of a potassium ion channel is an archetype for many ion-channel structures • ※Na+ channel • purified from the electric organ of electric eel. • bind a specific neurotoxin • (= Tetrodotoxin from puffer, 10ng lethal dose). • single 260kda chain. • contains 4 internal repeats(similar a.a. seq. gene duplication) • each repeat contains 5 hydrophobic segment(S1, S2,S3, S5, S6). • each repeat contains 1 highly positive charged segment(S4). • S1~S6 = membrane-spanning α helices. • S4 = voltage sensors of the channel.
※ K+ channel • 70 kDa protein that has 4 subunits(= tetramer). • S1~S6 = membrane-spanning α helices. • homologous to one of the repeated units of Na+ channels. • S5 and S6 = the actual pore of the K+ channel. • S1~S4 = apparatus that open the pore.
※ K+ channel (tetramer) Roderick MacKinnon determined the structure of a bacterial K+ channel in 1998 Nobel prize 2003
The structure of the potassium ion channel reveals the basis of ion specificity ※ K+ channel K+ ions must give up their water molecules and interact directly with groups from the protein. K+ Outside of the cell Pore becomes more constricted(3Å). K+ ion can fit in the pore without losing its shell of bound water(10Å). Inside of the cell
Selectivity filter : determines the preference for K+ over other ions (100-fold than Na+). • Conserved seq. : TVGYG extended confromation, • carbonyl oxgens line up.
K+ channels pass only K+. • Na+ is smaller than K+. • How is this high selectivity achieved?
Answer : Free energy costs of dehydrating these ions are considerable. • (Na+ =301kJ/mol, K+ =230kJ/mol) - The channel pays the cost of dehydrating K+ by providing compensating interactions with the carbonyl oxygen atoms lining the selectivity filter.
The structure of the potassium ion channel explains its rapid rate of transport High selectivity needs tight binding site! How to achieve fast transport of channels? Hydrated K+ ion proceeds into the channel. The ion then gives up its coordinated water molecules and binds to a site within the selectivity filter region The ion can move between the four sites(tetramer, similar ion affinity). This multiple binding site mechanism solves the apparent paradox of high ion selectivity and rapid flow.
Voltage gating requires substantial conformational changes in specific ion-channel domains • - S1~S4 forms domains termed “paddles”. • S4 = voltage sensor, highly positive charged residues.
Model for voltage gating of ion channels PADDLEs (S1-S4) include S4, the voltage sensor ※ Close state : paddles lie in a “down” position. ※ Depolarization : the paddles are pulled through the membrane into an “up” position. This motion pulls the base of the channel apart openining the channel
A channel can be inactivated by occlusion of the pore : The ball-and-chain model • Na+ and K+ channels undergo “inactivation” within miliseconds. • Cleavage of cytoplasmic side by trypsin → stayed persistently activation(channel always open). • Inactivation was restored by adding a synthetic peptide corresponding to the first 20 a.a cytoplasmic peptide Ball-and-stick model
- The first 20 residues of the K+ channel form a cytoplasmic unit(the ball) that is attached to a flexible segment of the polypeptide(the chain). - Channel is closed → the ball rotate freely. - Channel open → the ball quickly finds a complementary site in the open pore. And occludes it. Chain length & speed? Ball is postive charge?
The acetylcholine receptor is an archetype for ligand-gated ion channels • Neurotransmitter : small, diffusible signal molecules between a neuron and another neuron. • Ex> Acetylcholine • Synaptic cleft : a gap of about 50nm between presynaptic and postsynaptic membrane. • Nerve impulse → export of the contents of some 300 vesicles of acetylcholine into the cleft. • Acetylcholine receptor at postsynaptic membrane.
※Acetylcholine receptor • Ligand-gated channel. • Purified from electric ray. • 268kDa, pentamer of 4 kinds of membrane-spanning subunit(α2, β,γ, and δ). • Each subunit has a large extracellular domain + 4 hydrophobic segments(membrane spanning) + intracellular segment.
α2, β,γ, and δ - Pentamer, five-fold symmetry.
Binding of acetylcholine to the extracellular domain →structural alteration(rotations of the membrane spanning helices). - Large residues(leucine…) may occlude the channel by forming a tight hydrophobic ring. - The pore lined by small polar residues.
Action potentials integrate the activities of several ion channels working in concert + ions move to right side. + ions are accumulated at right side. Make electrostatic force. There are concentration gradients between two sides. Channel open Equilibrium Potential: concentration gradient is balanced by the electrostatic force resisting the motion of a additional charge.
※ Nernst equation Veq = -(RT/zF)ln([X]in/[X]out) = -(2.303)(RT/zF)log10([X]in/[X]out) R=gas constant F=Faraday constatn z=charge on the ion X • Membrane potential at equilibrium is called the equilibrium potential for a given ion at a given concentration ratio across a membrane. • [Na+]in= 14mM [Na+]out = 143mM, equilibrium potential = +62mV • [K+]in = 157mM [K+]out = 4mM, equilibrium potential = -98mV • The resting potential for a typical neuron is -60mV.
Acetylcholine binding Acetylcholine receptor channel open (Nonspecific cation channel) Na+ flow into / K+ flow out = -20mV Voltage gated Na+ channel open Na+ flow into rapidly = +30-mV Na+ channel inactivated Voltage gated K+ channel open K+ flow out slowly = -60mV K+ channel inactivated
13.5 Gap junctions allow ions and small molecules to flow between communicating cells • ※Gap junction • Cell-to cell channel,Passageways between the interiors of contiguous cells. • Clustered in discrete regions of the plasma membranes. • 20Å central hole. • The width of the gap between the cytoplasm ofthe two cells is about 35Å. • Small hydrophilic molecules( sugars, amino acids, nucleic acids, less than 1kda) can pass through gap junctions except protein, nucleic acid and polysaccharides. • Important for intercellular communication • insome excitable tissues, such as hear muscle.
A cell-to-cell channel is made of 12 molecules of connexin(30~42kda). • Connexin : 4 membrane spanning heices. • 6 connexin are hexagonally arrayed to form a half channel = connexon or hemichannle. • 2 connexons join end to end in the intercellular space. The closing of gap junctions by Ca2+ and H+ serves to seal normal cells from dying neighbors
13.6 Specific channels increase the permeability of some membranes to water • Some tissues need to transport water. • kidney, secretion of saliva and tears. • ※ Aquaporin (Peter Agre in red-blood-cell membrane) • Water channel. 24kda membrane protein • 6 membrane spanning helices. • Positive residues in the center prevent the transport of protons through aquaporin; maintain proton gradients