Membrane Channels
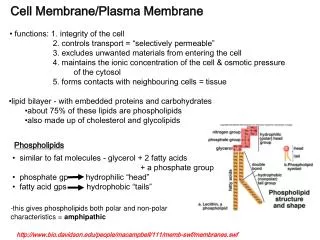
Membrane Channels. The Cell Membrane is Selective. Criteria for passage through the phospholipid bilayer : Hydrophobic Net zero charge Nonpolar Size is also a consideration Chemicals that will NOT pass through the phospholipid bilayer : Hydrophillic Charged, ionic Polar


Membrane Channels
E N D
Presentation Transcript
The Cell Membrane is Selective • Criteria for passage through the phospholipidbilayer: • Hydrophobic • Net zero charge • Nonpolar • Size is also a consideration • Chemicals that will NOT pass through the phospholipidbilayer: • Hydrophillic • Charged, ionic • Polar • Size is also a consideration
But, using the preceding criteria, many substances vital to cellular function (e.g., ions) and survival (e.g., glucose) will not gain entry into the cell! • So, nature has come up with channels, which selectively allow certain substances to gain entry into the cell, even though they do not meet the criteria on the preceding slide.
The Many Functions of Membrane Proteins Outside Plasma Membrane Inside Transported Enzyme Activity Cell Surface Receptor Cell Surface Identity Marker Cell Adhesion Attachment to the Cytoskleton
Channels are Vital Without channels it is energetically unfavorable to move ions across a membrane – 1. the phospholipidbilayer is ~6-8 nm thick. 2. the hydrophilic head of the phospholipid molecule projects toward the cytoplasm or the extracellular fluid. 3. the hydrophobic tails of the phospholipid molecules project toward each other.
Channel malfunction Cystic fibrosis Epilepsy Diabetes Migraines Neurotoxins Transmembrane Transport • a) communication among neurons • neural systems: • * action potential • * synaptic signaling • b) receptor – brain communication • heart muscle • signaling and regulatory processes
For the Cation to Move Through the PhospholipidBilayer…, • It must lose its waters of hydration so that it is not so huge and charged; requires energy to break attractive forces between the ion and the waters. • Energy is also required to move a charged highly hydrophilic particle into the highly hydrophobic area of the lipid bilayer that contains the “tails” of the phospholipid molecules. • Based on thermodynamic calculations, so much energy would be required for this process that it would never occur.
Channels are thermodynamically similar to Enzymes in that the former lower the Activation Energy required to move ions across the Membrane Transition State The rate of the reaction is determined by the energy of activation, the energy input required to produce the transition state. The uncatalyzed reaction requires a higher activation energy than the catalyzed one does. So, the latter runs more quickly. There is no difference in free energy (ΔG) between uncatalyzed and catalyzed reactions. The ΔG is the thermodynamic driving force for the reaction and determines the direction of the reaction. A + B AB C + D Reactants transition products (substrates) state
Ion Flow Across the Membrane • A Chemical can move across the membrane through one of two ways: • Movement through the phospholipidbilayer. • Movement through a H2O-filled protein channel.
Channels are thermodynamically similar to Enzymes in that the former lower the Activation Energy required to move ions across the Membrane The rate of diffusion is determined by the “energy of activation”, the energy input required to produce the “transition state”. (Remove H2Os of hydration and/or move into hydro-phobic environ-ment.) Movement through a channel only requires shedding of waters of hydration (energy input would be infinite to move through the bilayer). So. Diffusion occurs more quickly. ΔG determines this!! Transition State Change in free energy is the thermodynamic driving force for diffusion and determines the direction of ion movement A + B AB C + D Reactants transition products (substrates) state
What do we know about the structure of gated ion channels? • Biochemical Information – • MWs range from 25-250 kDal. • They are integral membrane glycoproteins. • They usually consist of 2 or more subunits. • The genes that code for the proteins have been isolated, cloned and sequenced. These sequences have been grouped into 6-7 protein families. • The primary (amino acid) sequences of these channels is known.
Use of the Hydrophobicity Plots • Propose 3-D structures of the channels • Propose functions for specific regions of the channel proteins
Amino Acid Sequence Enables Ion Channel Structure Determination
Many Techniques Can be Used to Test the Proposed Functions of Portions of the Channel Proteins • Sequence Homologies – used to determine which portions of primary sequences of ion channels are the same/very similar • Same channel from several different species – what is conserved must be critical to channel function e.g., Ach-gated channel – Ach receptor portion of the channel is highly conserved
Many Techniques Can be Used to Test the Proposed Functions of Portions of the Channel Proteins • Sequence Homologies – used to determine which portions of primary sequences of ion channels are the same/very similar (continued) b) Different channels with the same basic function from many tissues in one species – what is conserved must be critical to channel function e.g., Voltage gated channels (K+, Na+, Ca2+) – all have a presumed membrane-spanning region with charged AAs at each third position (voltage sensor?), while ligand-gated channels lack this structure
Many Techniques Can be Used to Test the Proposed Functions of Portions of the Channel Proteins 2) Immunocytochemistry– a) raise antibodies to a portion of the molecule thought to be on the intracellular or extracellular surface of the membrane b) Incubate neurons with the labeled antibodies c) Do antibodies bind to intact neurons? (Note: antibodies are too large to fit into a channel) Yes – sequence is on extracellular surface No – sequence may be in pore or on intracellular surface d) Next step: lyse cells and repeat exp. to see if there is binding, i.e. is it an intracellular sequence?
Many Techniques Can be Used to Test the Proposed Functions of Portions of the Channel Proteins 3) Site-directed mutagenesis – use molecular biology techniques to modify specific regions of a channel with a predicted function – does modification alter channel function in a predicted fashion? 4) Chimaeric Channel Construction – construct a mutant channel from sequences from 2 or more channel genes – which “parent” channel does the mutant resemble?
Most channels have this basic structure: multimeric (quarternary structure), membrane-spanning, and, by definition, have a pore running longitudinally through the structure. Vary in the number of subunits and complexity.
Remember your amino acids? • Primary, secondary, and tertiary structures of proteins. • In addition, recall that multimeric proteins are formed from the attraction of individual subunits, forming the quarternary structure. • Recall the structure and ionization of the each of the amino acid side-chains (R). -It wouldn’t hurt if you reviewed what a pI is.
The amino acid side –chains (R)
The primary amino acid sequence and higher –order structures determine the channel topology. Interior of the channel will be lined with hydrophilic amino acids. Exterior of the channel will be lined with hydrophobic amino acids.
Selectivity Filter • Many channels are selective for only 1 or 2 different chemicals (ions, sugars, etc.). • The K+ channel has such a filter, which is a narrow region towards the extracellular surface of the membrane. • Two K+ ions can occupy the selectivity filter simultaneously, with a third in a H2O-filled cavity deeper in the pore.
Proposed Mechanisms for Channel Ion Selectivity Non-specific cation channel, i.e. little selectivity other than for cations 10-20 X more Na+ than K+ 100 X more K+ than Na+ Voltage-gated K+ channel – 3.3 A in diameter Voltage-gated Na+ channel - 4 A in diameter Ach receptor channel - 6.5 A in diameter
Proposed Mechanisms for Channel Ion Selectivity by Channels: Ionic size Non-specific cation channel, i.e. little selectivity other than for cations 10-20 X more Na+ than K+ 100 X more K+ than Na+ Ach receptor channel - 6.5 A in diameter Voltage-gated K+ channel – 3.3 A in diameter Voltage-gated Na+ channel - 4 A in diameter Non-hydrated Na+ ion = 1.9 A in diameter Non-hydrated K+ ion = 2.7 A in diameter If ionic size explains channel selectivity, why is the K+ channel so selective for K+ since Na+ is smaller?
Proposed Mechanisms for Ion Selectivity by Channels: Ionic size Non-specific cation channel, i.e. little selectivity other than for cations 10-20 X more Na+ than K+ 100 X more K+ than Na+ Ach receptor channel - 6.5 A in diameter Voltage-gated K+ channel – 3.3 A in diameter Voltage-gated Na+ channel - 4 A in diameter Hydrated Na+ ion = 3.3-4 A in diameter Modified Model = perhaps channels select based on hydrated ionic radius? Hydrated K+ ion = 3.3 A in diameter (K+ is larger, has a lower charge density and so attracts fewer waters of hydration.)
Proposed Mechanisms for Ion Selectivity by Channels: Ionic size The modified model explains K+ channel selectivity, i.e. the hydrated K+ just fits into the channel and the hydrated Na+ is too big to fit. However, how do we explain the +/- sodium channel selectivity? A selectivity filter exists inside the channel
Proposed Mechanisms for Ion Selectivity by Channels: Ionic size How might it work? Similar to enzymes, but much faster? Sodium recognition site = selectivity filter Na+ Na+
Evidence for a Selectivity Filter If channels are simple resistors, than movement through an open channel should be a function of the concentration gradient for the ion across the membrane Rate of ion movement = ax[ion]I/[ion]o (current flow = diffusion) Linear relationship with slope = a
Evidence for a Selectivity Filter Observed data for Na+ channel Unitary current (pa) = recordings from single channels Expected data External [Na+] mM
Evidence for a Selectivity Filter Data for voltage-gated Na+ channel do not fit the model of a channel as a simple resistor in the membrane. Instead, the current flow through the Na+ channel plateaus or “saturates” at high [Na+]. This relationship looks like what happens to an enzyme at high [substrate]. Perhaps some channels select ions based on the same biochemical mechanisms used by enzymes to select their substrates? In the end, the final determinations of channel gating mechanisms and ion selectivities will come from X-ray crystallography of the purified channels.
Crystal Structure of the K+ Channel from above and from the side The K+ channel is structure such that a very narrow tube through the inverted cone shape allows for only 50 H2O molecules and only 2 K+ in succession. Because they strongly repel each other, when one enters, one will be forced out.
Methods for Studying Ion Channels - 1 Biochemistry • agonist, antagonist or drug binding • isolation and purification • reconstitution Molecular biology • sequencing, cloning, mutagenesis Structural biology • microscopy, crystallography, NMR, ...
Methods for Studying Ion Channels - 2 Electrophysiology • tissue slice • extracellular recording • intracellular recording • whole-cell recording • single channel recording Biochemistry • radioactive ion flux Voltage clamp Current clamp Concentration jump
Cloning via Protein Purification Pure Na channel proteins Electric eel Protein Purification Na channel proteins Microsequence Oligonucleotide probe with sequence corresponding to aa sequence Design probe Amino acid sequence of small region of Na channel protein Hybridize to cDNA library containing Na channel cDNA Isolate and sequence Na channel cDNA AA sequence of entire Na channel protein s s s s s s s s s s s s s s s s Synthesize cDNA library containing Na channel cDNA (s) Deduce protein sequence sss sss sss Isolate mRNAs including one encoding Na channel
Positional cloning – Shaker flies • Cloning by sequence homology - Use the same strategy as that used in preceding slide, except that the sequence on which the cloning is based comes not from the purified protein, but rather from the already-isolated cDNA. Normal Shaker
Positional Cloning Enabled Determination of the Shaker K Channel
Typical Ion Channels with Known Structure: Acetylcholine receptor M2 transmembrane segment K+ channel (KCSA) • Types of ion channels: • Simple pores (GA, GAP junctions) • Substrate gated channels (Nicotinic receptor) • Voltage-gated channels (K-channels) • Pumps (ATP-synthase, K+,Na+-ATPase)
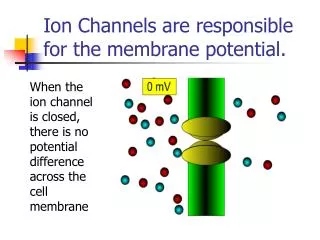
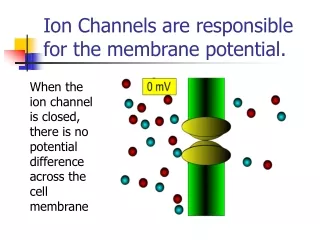
ion channels in the PM of neurons and muscles contributes to their excitability • when open - ions move down their concentration gradients • channels possess gates to open and close them • two types: gated and non-gated Ion Channels • 1. Leakage (non-gated) or Resting channels: are always open, contribute to the resting potential • -nerve cells have more K+ than Na+ leakage channels • -as a result, membrane permeability to K+ is higher • -K+ leaks out of cell - inside becomes more negative • -K+ is then pumped back in • 2. Gated channels: open and close in response to a stimulus • A. voltage-gated: open in response to change in voltage - participate in the AP • B. ligand-gated: open & close in response to particular chemical stimuli (hormone, neurotransmitter, ion) • C. mechanically-gated: open with mechanical stimulation
Types of Biochemical Mechanisms that Open and Close Channels (Cont’d) • Nt or hormone binding to receptor causes a 2nd messenger to activate a protein kinase that phosphorylates a channel and thus opens it. • Changes in membrane potential. • Membrane deformation (e.g., mechanical pressure). • Selectivity by charge (i.e., positively lined pore allows anions through; negatively lined pore allows cations through).
Na+ Channels have Gates At rest, one is closed (the activation gate) and the other is open (the inactivation gate). Suprathreshold depolarization affects both of them.
The resting potential, recall, is generated mainly by open “resting”, non-gated K+ channels -the number of K+ channels dramatically outnumbers that of Na+ -however, there are a few Na leak channels along the axonal membrane ECF AXON
Channel Gating Mechanisms AChR: Proposed gating mechanism (Unwin, 1995) Open Closed
Channel Families • Voltage-gated • Extracellular ligand-gated • Intracellular ligand-gated • Inward rectifier • Intercellular • Other
Voltage-gated • sodium: I, II, III, µ1, H1, PN3 • potassium: KA, Kv (1-5), Kv(r), Kv(s),KSR, BKCa, IKCa, SKCa, KM, KACh • calcium: L, N, P, Q, T • chloride: ClC-0 - ClC-8
Extracellular ligand-gated • nicotinic ACh (muscle): 2 (embryonic), 2 (adult) • nicotinic ACh (neuronal): (2-10), (2-4) • glutamate: NMDA, kainate, AMPA • P2X (ATP) • 5-HT3 • GABAA: (1-6), (1-4), (1-4), , , (1-3) • Glycine
Intracellular ligand-gated • leukotriene C4-gated Ca2+ • ryanodine receptor Ca2+ • IP3-gated Ca2+ • IP4-gated Ca2+ • Ca2+-gated K+ • Ca2+-gated non-selective cation • Ca2+-gated Cl– • cAMP cation • cGMP cation • cAMP chloride • ATP Cl– • volume-regulated Cl– • arachidonic acid-activated K+ • Na+-gated K+
Inward rectifier • Kir • 1.1-1.3 • 2.1-2.4 • 3.1-3.5 • 4.1-4.2 • 5.1 • 6.1-6.2 • 7.1