Download

1 / 22
410 likes | 2.5k Vues
CELLULAR BIOCHEMISTRY PROTEINS AND ENZYMES - LECTURE 8 ENZYMES - MECHANISM OF ACTION - LYSOZYME. FACTORS THAT ENHANCE REACTION RATE LYSOZYME MECHANISM. MECHANISMS OF ENZYME ACTION - GENERAL ASPECTS. FACTORS RESPONSIBLE FOR ENZYMIC CATALYSIS: 1. Proximity and orientation of substrate

E N D
CELLULAR BIOCHEMISTRY PROTEINS AND ENZYMES - LECTURE 8 ENZYMES - MECHANISM OF ACTION - LYSOZYME FACTORS THAT ENHANCE REACTION RATE LYSOZYME MECHANISM
MECHANISMS OF ENZYME ACTION - GENERAL ASPECTS FACTORS RESPONSIBLE FOR ENZYMIC CATALYSIS: 1. Proximity and orientation of substrate 2. Formation of specific, unstable covalent intermediates 3. Acid-base catalysis 4. Strain and stabilisation of transition state intermediates 5. Induced fit 6.Hydrophobic environment For any given enzyme several of the above factors will apply and the enhanced reaction rate is therefore the productof several separate enhancements. Further reading: Stryer 5th edn. Chapters 8 and 9. For lysozyme mechanism of action - Stryer 4th Edn, pp.207-216 gives a full and clear account
Proximity and orientation of substrate • Binding of two or more substrates together at the active site increases their effective concentrations. • Specific binding at the active site can orientate the substrates precisely with regard to each other and the catalytic groups on the enzyme. Koshland has suggested that such orientation may involve orbital steering, the electron shells being brought into very precise proximity. Note that these factors contribute to specificity as well as catalysis.
Formation of specific, unstable covalent intermediates Chymotrypsin, acting as an esterase, cleaves p-nitrophenyl acetate to give a yellow product, p-nitrophenol. The reaction goes in two stages: chymotrypsin + p-nitrophenyl acetate ———> acetyl chymotrypsin + p-nitrophenol fast acetyl chymotrypsin + water ———> chymotrypsin + acetate slow By lowering the pH rapidly, the acetyl chymotrypsin intermediate can be isolated. The acetyl is attached covalently to a serine side-chain at the active site.
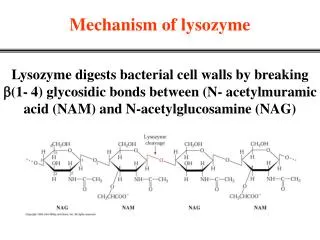
Acid-base catalysis In the chemistry laboratory acid-base catalysis is achieved at extremes of pH. Enzymes achieve general acid-base catalysis at near neutral pH by using amino acid side chains that can act as as proton donors (acids) or proton acceptors (bases). For example in lysozyme a glutamic acid side chain [-(CH2)2-COOH] donates a proton to the susceptible bond in the polysaccharide substrate, causing cleavage. Two side chain groups that can act as proton donors: Two side chain groups that can act as proton acceptors:
Strain and stabilisation of transition state intermediates • Often when the substrate binds to its active site, part of its conformation is forced to change in order for it to 'fit’. The new conformation favours formation of transition stateintermediates (i.e. such intermediates are stabilised by the geometry of the active site) and destabilises the substrate. • The reaction is therefore facilitated. Note that the strain imparted to the conformation of the substrate is a consequence of good binding of the enzyme to the rest of the substrate (high affinity). • Again lysozyme is a good example; as the polysaccharide substrate binds to the active site it mostly fits well (facilitating binding) but the sugar residue next to the cleavable bond is distorted, destabilizing the substrate, but stabilizing the transition state intermediate.
Induced fit Koshland's theory. The conformation of the enzyme may change as the substrate binds, contributing to the overall strain in the E-S complex. Also, induced fit may play an important role in specificity as well as catalytic activity. It can explain why an enzyme like hexokinase can phosphorylate sugars, but not H2O. Hexokinase catalyses: glucose + ATP ———> glucose-6-phosphate + ADP but not H2O + ATP ———> phosphate + ADP (the second reaction would waste ATP, and has to be avoided) Induced fit here means that the enzyme's conformationchanges (and the active site becomes active) only when glucose is bound to the active site. With just water in the active site, the conformation is the inactive one, and the water cannot be phosphorylated.
CONFORMATIONAL CHANGE OF HEXOKINASE (From Stryer Ch 16)
Hydrophobic environment • Water is often a poor solvent in which to carry out chemical reactions because the reactive groups become solvated (hydrated) and surrounded by a shell of water molecules that prevents the approach of another reactant. • Organic chemists usually carry out their reactions in organic solvents which are less effective in shielding groups in this way. • The active site of an enzyme often contains a number of hydrophobic side chains and, as the substrate binds, water molecules are squeezed out and a hydrophobic micro-environment is established in which hydration/shielding of reactive groups is minimised. • So the environment in the immediate region of the reactive groups resembles that favoured by the organic chemist.
LYSOZYME ACTION WHICH BOND IS CLEAVED? 1. Rate of hydrolysis of oligomers of NAG (relative): (NAG)20 (NAG)3 1 (NAG)48 (NAG)54,000 (NAG)630,000 (NAG)830,000 2. (NAG)3 is stable, so probably A-B and B-C bonds are not cleaved 3. Site C on enzyme cannot be filled by NAM (lactyl side-chain will not fit), so bacterial substrate must fit: NAG-NAM-NAG-NAM-NAG-NAM A B C D E F Lysozyme specifically cleaves a NAM-NAG bond, so C-D and E-F cannot be cleavage sites. By elimination cleavage site must be between D and E
LYSOZYME ACTION WHICH SIDE OF D-E BOND IS CLEAVED? Distinguished by use of water enriched with 18O Skeleton structures only. So, cleavage must be at position a
MECHANISM OF ACTION OF LYSOZYME Cleavage of(NAG)6: NAG-NAG-NAG-NAG-NAG-NAG A B C D E F 1. -COOH of Glu-35 donates H+ to bond between C1 of D ring and glycosidic oxygen, cleaving bond: 2. This creates a positive charge on C1 of D NAG residue - a transient intermediate carbonium ion. Potentially very unstable, but stabilised by distortion of D-ring and proximity of negative charge on Asp-52.
MECHANISM OF ACTION OF LYSOZYME (CONT) Cleavage of(NAG)6: NAG-NAG-NAG-NAG-NAG-NAG A B C D E F 3. Dimer (residues E-F) diffuses away 4. Carbonium ion intermediate reacts with OH- from solvent:: 5. (NAG)4 diffuses away. Glu-35 is reprotonated.
MECHANISM OF ACTION OF LYSOZYME (CONT) CHAIR AND HALF-CHAIR CONFORMATIONS
MECHANISM OF ACTION OF LYSOZYME (CONT) pH PROFILE
MECHANISM OF ACTION OF LYSOZYME (CONT) TRANSITION STATE ANALOGUE The lactone analogue resembles the transition-state intermediate because its D ring has a half-chair form
MECHANISM OF ACTION OF LYSOZYME Contributions of each sugar of (NAG)6to standard free energy of binding of substrate