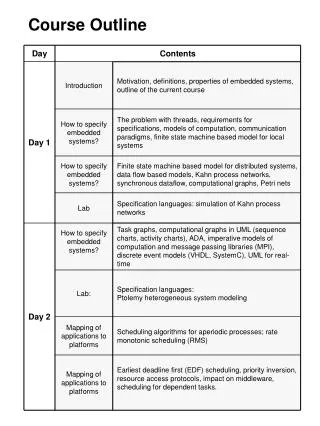
Course outline
Course outline. Membrane transport. Cell membranes. www.cellsalive.com/. Cell membranes. Cell membranes. At very high magnification & in color. Membrane structure. http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/C/CellMembranes.html. Cell membranes.


Course outline
E N D
Presentation Transcript
Cell membranes www.cellsalive.com/
Cell membranes At very high magnification & in color
Membrane structure http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/C/CellMembranes.html
Cell membranes Every cell is encircled by a membrane and most cells contain an extensive intracellular membrane system. Membranes fence off the cell's interior from its surroundings. Membranes let in water, certain ions and substrates and they excrete waste substances. They act to protect the cell. Without a membrane the cell contents would diffuse into the surroundings, information containing molecules would be lost and many metabolic pathways would cease to work. The cell would die!
Cell membranes • Surround all cells • Fluid-like composition, like soap bubbles • Composed of: • Lipids in a bilayer • Proteins embedded in lipid layer (called trans-membrane proteins) • And, Proteins floating within the lipid sea (called integral proteins) • And Proteins associated outside the lipid bi-layer (peripheral).
Membrane transport • Transporters are of two general classes: • carriers and channels. • These are exemplified by two ionophores (ion carriers produced by microorganisms): • valinomycin (a carrier) • gramicidin (a channel).
Valinomycin H C C H 3 3 C H O O O C H O 3 H H N C H C O C N C O C H C C C H C H H C H 3 H C C H H C C H 3 3 3 3 - valine - hydroxy - - valine - lactic L D D L isovaleric acid acid Valinomycin is a carrier for K+. It is a circular molecule, made up of 3 repeats of the sequence shown above.
Valinomycin Valinomycin O O O + K O O O Hydrophobic Valinomycin reversibly binds a single K+ ion. The ring closely surrounds the K+ ion, which interacts with 6 oxygen atoms of valinomycin. Valinomycin is highly selective for K+ relative to Na+. The smaller Na+ ion cannot simultaneously interact with all 6 oxygen atoms within valinomycin. Thus it is energetically less favorable for Na+ to shed its water of hydration to form a complex with this ionophore.
Valinomycin Valinomycin O O O + K O O O Hydrophobic Whereas the interior of the valinomycin-K+ complex is polar, the surface of the complex is hydrophobic. Valinomycin enters the lipid core of the bilayer and solubilizes K+ within this hydrophobic milieu. Crystal structure (at Virtual Museum of Minerals & Molecules).
Valinomycin + + Val - K Val - K + + K K Val Val membrane Valinomycin is a passive carrier for K+. It can bind or release K+ when it encounters the membrane surface. Valinomycin can catalyze net K+ transport because it can translocated either in the complexed or uncomplexed state. The direction of net flux depends on the electrochemical K+ gradient.
Proteins as carrier • Proteins that act as carriers are too large to move across the membrane. • They are transmembrane proteins, with fixed topology. • Example: GLUT1 glucose carrier, found in plasma membranes of various cells, including erythrocytes. • GLUT1 is a large integral protein, predicted via hydropathy plots to include 12 transmembrane α-helices.
Proteins as carrier conformation conformation change change Carrier mediated solute transport Carrier proteinscyclebetweenconformations in which a solute binding site is accessible on one side of the membrane or the other. There may be an intermediate conformation in which a bound substrate is inaccessible to either aqueous phase. With carrier proteins, there is never an open channel all the way through the membrane.
Kinetics of transport carriers Carriers exhibit Michaelis-Mentenkinetics. The transport rate mediated by carriers is faster than in the absence of a catalyst, but slower than with channels. A carrier transports only one or few solute molecules per conformational cycle.
Classes of carrier proteins Uniport Symport Antiport A A B A B Uniport (facilitated diffusion) carriers mediate transport of a single solute. Examples include GLUT1 and valinomycin. These carriers can undergo the conformational change associated with solute transfer either empty or with bound substrate. Thus they can mediate net solute transport.
Classes of carrier proteins Uniport Symport Antiport A A B A B Symport(cotransport) carriers bind 2 dissimilar solutes (substrates) & transport them together across a membrane. Transport of the 2 solutes is obligatorily coupled. A gradient of one substrate, usually an ion, may drive uphill (against the gradient) transport of a co-substrate. An example is the plasma membrane glucose-Na+ symport.
Classes of carrier proteins Uniport Symport Antiport A A B A B Antiport (exchange diffusion) carriers exchange one solute for another across a membrane. Usually antiporters exhibit "ping pong" kinetics. One substrate is transported across a membrane and then another is carried back. Example: ADP/ATP exchanger (adenine nucleotide translocase) which catalyzes 1:1 exchange of ADP for ATP across the inner mitochondrial membrane.
Active transport Active ADP + Pi Transport S S 2 1 ATP Side 1 Side 2 Active transport enzymes couple net solute movement across a membrane to ATP hydrolysis. An active transport pump may be a uniporter, or it may be an antiporter that catalyzes ATP-dependent transport of 2 solutes in opposite directions. ATP-dependent ion pumps are grouped into classes, based on transport mechanism, genetic & structural homology.
Ion pumps P-class ion pumpsare a gene family exhibiting sequence homology. They include: • Na+,K+-ATPase, in plasma membranes of most animal cells, is an antiport pump. It catalyzes ATP-dependent transport of Na+ out of a cell in exchange for K+ entering. • (H+, K+)-ATPase, involved in acid secretion in the stomach, is an antiport pump. It catalyzes transport of H+ out of the gastric parietal cell (toward the stomach lumen) in exchange for K+ entering the cell.
Ion pumps P-class pumps (cont): • Ca++-ATPases, in endoplasmic reticulum (ER) & plasma membranes catalyze transport of Ca++ away from the cytosol, either into the ER lumen or out of the cell. There is some evidence that H+ may be transported in the opposite direction. Ca++-ATPase pumps keep cytosolic Ca++ low, allowing Ca++ to serve as a signal.
Ion pumps O E n z y m e - C O H P A T P i A D P H O 2 O O E n z y m e - C O P O - O - P - Class Pumps The reaction mechanism for a P-class ion pump involves transient co-valent modification of the enzyme. At one stage of the reaction cycle, Pi is transferred from ATP to the carboxyl of a Glu or Asp residue, forming a “high energy” anhydride linkage (~P). At a later stage in the reaction cycle, the phosphate is released by hydrolysis.
Ca++ pump ++ ++ ~ ~ E P - Ca E P - Ca 2 2 ADP ++ 2Ca ATP ++ E - Ca 2 ++ 2Ca E P i ER cytosol membrane lumen The ER Ca++ pump is called SERCA: Sarco(Endo)plasmic Reticulum Ca++-ATPase. In this diagram of the SERCA reaction cycle, conformational changes altering accessibility of Ca++-binding sites to the cytosol or ER lumen are depicted as positional changes. Keep in mind that SERCA is a large protein that maintains its transmembrane orientation.
Ca++ pump ++ ++ ~ ~ E P - Ca E P - Ca 2 2 ADP ++ 2Ca ATP ++ E - Ca 2 ++ 2Ca E P i ER cytosol membrane lumen Reaction cycle 1 2 Ca++ bind tightly from the cytosolic side, stabilizing the conformation that allows ATP to react with an active site aspartate residue. 2 Phosphorylation of the active site aspartate induces a conformational change that shifts accessibility of the 2 Ca++ binding sites from one side of the membrane to the other, & lowers the affinity of the binding sites for Ca++.
Ca++ pump ++ ++ ~ ~ E P - Ca E P - Ca 2 2 ADP ++ 2Ca ATP ++ E - Ca 2 ++ 2Ca E P i ER cytosol membrane lumen 3Ca++ dissociates into the ER lumen. 4Ca++ dissociation promotes hydrolysis of Pi from the enzyme Asp and the conformational change (recovery) that causes the Ca++ binding sites to be accessible again from the cytosol.
SERCA structure Asp351 cytosolic domain membrane 2 Ca++ domain Muscle SERCA PDB 1EUL The structure of muscle SERCA, determined by X-ray crystallography, shows 2Ca++ ions bound between transmembrane α-helices. These intramembrane Ca++ binding sites are presumed to participate in Ca++ transfer across the membrane.
SERCA structure Asp351 cytosolic domain membrane 2 Ca++ domain Muscle SERCA PDB 1EUL The active site Asp351, which is transiently phosphorylated during catalysis, is in a cytosolic domain, far from the Ca++ binding sites. The sequence adjacent to Asp351 (DKTGTLT) is in all P-class pumps. Ca++ has been found to induce large structural changes in cytosolic and transmembrane domains of SERCA, consistent with the proposed conformational coupling between active site and membrane domains.
Ca++ transport Ca++ SERCA Conformational Cycle enzyme phosphate phosphorylation hydrolysis Observed changes in rotation and tilt of transmembrane a-helices may be involved in altering access of Ca++ binding sites to one side of the membrane or the other, and altering the affinity of binding sites for Ca++, at different stages of the SERCA reaction cycle. Only 2 transmembrane a-helices are represented above. Animation of mechanism by MacLennan lab.
Trans-epithelial transport glucose Na+ glucose - Na+ symport apical end glucose Na+ ATP ADP + P i basal end Na+ pump GLUT2 K+ intestinal epithelial cell In the example shown, 3 carrier proteins accomplish absorption of glucose & Na+ in the small intestine. This transport across a cell layer depends on localization of specific plasma membrane transporters at either the apical end of each epithelial cell (facing the intestinal lumen) or the basal end (facing a blood capillary).
Trans-epithelial transport glucose Na+ glucose - Na+ symport apical end glucose Na+ ATP ADP + P i basal end Na+ pump GLUT2 K+ intestinal epithelial cell • The Na+ pump, at the basal end of the cell, keeps [Na+] lower in the cell than in fluid bathing the apical surface. • The Na+ gradient drives uphill transport of glucose into the cell at the apical end, via glucose-Na+ symport. [Glucose] within the cell is thus higher than outside. • Glucose flows passively out of the cell at the basal end, down its gradient, via GLUT2 (uniport related to GLUT1).
Ion channels conformation change open closed Channels cycle between open & closed conformations. When open,a channel provides a continuous pathway through the bilayer. Whereas carriers transport only one or a few ions or molecules per conformational cycle, many ions flow through a channel, each time it opens. Transport rates are higher for channels than for carriers.
Gating Proposed mechanism of gramicidin gating closed open Gating (opening & closing) of a gramicidin channel is thought to involve reversible dimerization. An open channel forms when two gramicidin molecules join end to end to span the membrane. This model is consistent with the finding that at high [gramicidin] overall transport rate depends on [gramicidin]2.
A look at history Since the origins, Computer Scientist have looked to relationships among machines and living organisms • McCulloch and Pitts, Neural Networks, 1943 • Von Neumann, Cellular Automata, 1966 • Lindenmayer, L systems, 1968 • Holland, Genetic Programming, 1975
L-systems L-systems are a mathematical formalism proposed by the biologist Aristid Lindenmayer in 1968 as a foundation for an axiomatic theory of biological development. More recently, L-systems have found several applications in computer graphics. Two principal areas include generation of fractals and realistic modelling of plants
Molecular computing “....Theoretical arguments suggest that more efficient and adaptable modes of computing are possible, while emerging biotechnologies point out to possibilities for implementation. Their common ground is molecular computing... It is likely that molecular computing will prove more valuable outside the context of conventional Von Neumann computers. Critically important computing needs such as adaptive patterns and process control may be refractory to simple decreases in size and increases in speed. Instead of suppressing the unique properties of carbon polymers, we should consider how to harness them to fill these needs....” Michael Conrad, On Design Principles for a Molecular Computer, 1985
Another quote ....Inheritance is a discourse, a set of instructions passed from generation to generation. It has a vocabulary -the genes themselves- a grammar, the way in which the information is arranged, and a literature, the thousands of instructions needed to make a human being...” Steve Jones, The Language of The Genes, 1993
How cells process information • DNA may be viewed as a double sequence of four symbols: A, T, C, G • DNA is naturally processed by duplication, recombination, etc. • Biologist and Genetists have developed so far a variety of techniques to manipulate DNA sequences (Biotechnologies) • Information stored in DNA sequences is translated into proteins by DNA Transcription • Proteins control and regulate the activity of the genes (Gene Expression)
Membrane computing • Gh. Păun, Computing with Membranes, 1998 • Membrane Computing looks at the whole cell structure and functioning as a computing device • Membranes play a fundamental role in the cell as filters and separators • Modeling the living cell is beyond the purpose of Membrane Computing
references • Păun, Gh., Membrane Computing. An Introduction, Springer-Verlag, Berlin, 2002. • Păun, Gh., Rozenberg, G., Salomaa, A., Zandron, C. (eds.), Membrane Computing, LNCS, 2597, Springer-Verlag, 2003. • Cavaliere, M., Martin-Vide, C., Paun, Gh. (eds.), Brainstorming Week on Membrane Computing, Technical Report of the Research Group on Mathematical Linguistics, N. 26/03, Universitat Rovira I Virgili, Tarragona, Spain, 2003. • The P systems Web Pages, http://psystems.disco.unimib.it • Alberts, B., et al., Molecular Biology of the Cell, Garland Science, New York, 2002.
A membrane system (or P-system) • A membrane structure formed by several membranes embedded in a unique main membrane • Multi-sets of objects placed inside the regions delimited by the membranes (one per each region) • The objects are represented as symbols of a given alphabet (each symbol denotes a different object) • Sets of evolution rules associated with the regions (one per each region), which allow the system • to produce new objects starting form the existing ones • to move objects from one region to another
A membrane system (or P-system) • Each region contains a multi-set of objects and a set of rules. The objects are represented by symbols from a given alphabet. Typically, a evolution rule from region r is of the form ca→cbindoutdhere and it says that a copy of object a in the presence of a copy of the catalyst c is replaced by a copy of the object b and 2 copies of the object d. • b has to immediately enter the inner membrane of region r labeled j, a copy of d is sent out through the membrane of region r and a copy of d remains in r.
A computation in a P-system • We start with an initial configuration: an initial membrane structure and some initial multi-sets of objects placed inside the regions of the system. • We apply the rules in a non-deterministic maximal parallel manner: in each step, in each region, each object that can be evolved according to some rule must do it • A computation is said successful if it halts, that is, it reaches a configuration where no rules can be applied. • The result of a successful computation may be the multi-sets formed either by the objects contained in a specific output membrane or by the objects sent out of the systems during the computation • A non-halting computation yields no result
An other example R1: aa → (a,here)(a,in),ab → (b,here)(a,in) R2: a → (a,out)(b,out)(b,in),a → (c,in) R3: b → (a,here)(a,out)(b,in),cb → (a,here) R4: Ø