Download
1 / 85
970 likes | 1.91k Vues
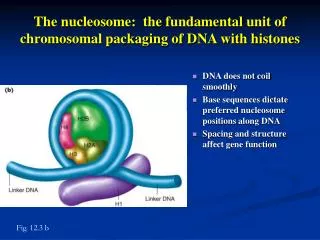
The nucleosome: the fundamental unit of chromosomal packaging of DNA with histones. DNA does not coil smoothly Base sequences dictate preferred nucleosome positions along DNA Spacing and structure affect gene function. Fig. 12.3 b.

E N D
The nucleosome: the fundamental unit of chromosomal packaging of DNA with histones • DNA does not coil smoothly • Base sequences dictate preferred nucleosome positions along DNA • Spacing and structure affect gene function Fig. 12.3 b
Models of higher level compaction seek to explain extreme compaction of chromosomes at mitosis • Formation of 300 A fiber through supercoiling Fig. 12.4 a
Models of higher level compaction seek to explain extreme compaction of chromosomes at mitosis • Radial loop-scaffold model for higher levels of compaction • Each loop contains 60-100 kb of DNA tethered by nonhistone scaffold proteins Fig. 12.4 b
Radial loop-scaffold model continued Fig. 12.4 c
A closer look at karyotypes: fully compacted metaphase chromosomes have unique, reproducible banding patterns • Banding patterns are highly reproducible • Not known what they represent Fig. 12.6 a
A closer look at karyotypes • Banding patterns help locate genes Fig. 12.6 b
Polytene chromosomes are an invaluable tool for geneticists • in situ hybridization of white gene to a single band (3C2) near the tip of the Drosophila X chromosome Fig. 12.15 c
A closer look at karyotypes • Banding patterns can be used to analyze chromosomal differences between species • Can also be used to reveal cause of genetic disease • e.g., Downs syndrome – 3 copies of chromosome 21 Fig. 12.6 c
Specialized chromosomal elements ensure accurate replication and segregation of chromosomes • There are many origins of replication • Replication occurs in about 8 hours during S phase in actively dividing human cells • DNA polymerase can assemble new DNA at a rate of about 50 nucleotides per second • Many origins of replication are required to complete the task of copying the DNA in a genome • In mammals, there are 10,000 origins of replication • Origins of replication are scattered throughout the chromatin, 30 – 300 kb apart
Structure of yeast origin of replication • Autonomously replicating sequences (ARSs) in yeast consist of an A – T rich region • ARSs permit replication of plasmids in yeast cells Fig. 12.11 b
Telomeres preserve the integrity of linear chromosomes • Telomeres are protective caps on eukaryotic chromosomes • Prevent fusion with other chromosomes • Protect tips from degradation • Solve the end-replication problem Fig. 12.8
DNA polymerase cannot reconstruct 5’ end of a DNA strand Fig. 12.9
Binding of telomerase to TTAGGG and addition of RNA extends the ends Fig. 12.10
Segregation of condensed chromosomes depends on centromeres • Centromeres appear as constrictions on chromosomes • Contained within blocks of repetitive, noncoding sequences called satellite DNA • Satellite DNA consists of short sequences 5-300 bases in length • Centromeres have two functions • Hold sister chromatids together • Kinetochore – structure composed of DNA and protein that help power chromosome movement
Centromere structure and function Fig. 12.11 a
Structure of yeast centromere Fig. 12.11 b
Studies using DNase identify decompacted regions Fig. 12.12 a
Position effect variegation in Drosophila: moving a gene near heterochromatin prevents it expression • Facultative heterochromatin • Moving a gene near heterochromatin silences its activity in some cells and not others Fig. 12.14 a
The loss or gain of one or more chromosomes results in aneuploidy
Autosomal aneuploidy is harmful to the organism • Monosomy usually lethal • Trisomies – highly deleterious • Trisomy 18 – Edwards syndrome • Trisomy 13 – Patau syndrome • Trisomy 21 – Down syndrome
Humans tolerate X chromosome aneuploidy because X inactivation compensates for dosage Fig. 13.27
Meiotic nondisjunction • Failure of two sister chromatids to separate during meiotic anaphase • Generates reciprocal trisomic and monosomic daughter cells • Chromosome loss • Produces one monosomic and one diploid daughter cell Fig. 13.28 a
Mosaics – aneuploid and normal tissues that lie side-by-side • Aneuploids give rise to aneuploid clones Fig. 13.28 b
Gynandromorph in Drosophila results from female losing one X chromosome during first mitotic division after fertilization Fig. 13.29
Euploid individuals contain only complete sets of chromosomes
Monoploid organisms contain a single copy of each chromosome and are usually infertile • Monoploid plants have many uses • Visualize recessive traits directly • Introduction of mutations into individual cells • Select for desirable phenotpyes (herbicide resistance) • Hormone treatment to grow selected cells
Treatment with colchicine converts back to diploid plants that express desired phenotypes Fig. 13.30 c
Polyploidy has accompanied the evolution of many cultivated plants • 1 out of 3 flowering plants are polyploid • Polyploid often increases size and vigor • Often selected for agricultural cultivation • Tetraploids - alfalfa, coffee, peanuts • Octaploid - strawberries Fig. 13.31
Triploids are almost always sterile • Result from union of monoploid and diploid gametes • Meiosis produces unbalanced gametes Fig. 13.32
Tetraploids are often source of new species • Failure of chromosomes to separate into two daughter cells during mitosis in diploid • Cross between tetraploid and diploid creates triploids – new species, autopolyploids 13.33 a
Maintenance of tetraploid species depends on the production of gametes with balanced sets of chromosomes • Bivalents- pairs of synapsed homologous chromosomes that ensure balanced gametes Fig. 13.33 b
Some polyploids have agriculturally desirable traits derived from two species • Amphidiploids created by chromosome doubling in germ cells • e.g., wheat – cross between tetraploid wheat and diploid rye produce hybrids with desirable traits Fig. 13.34
Phenotypic consequences of heterozygosity • Homozygosity for deletion is often but not always lethal • Heterozygosity for deletion is often detrimental Fig. 13.3
Mapping distances affected in deletion heterozygotes • Recombination between homologues can only occur if both carry copies of the gene • Deletion loop formed if heterozygous for deletion • Genes within the loop cannot be separated by recombination Fig. 13.4 a
Deletion loops in polytene chromosomes Fig. 13.4 b
Deletions in heterozygotes can uncover genes • Pseudodominance shows a deletion has removed a particular gene Fig. 13.5
Deletions can be used to locate genes • Deletions to assign genes to bands on Drosophila polytene chromosomes • Complementation tests • Deletion heterozygote reveals chromosomal location of mutant gene Fig. 13.6
Deletions to locate genes at the molecular level • Labeled probe hybridizes to wild-type chromosome but not to deletion chromosome Fig. 13.7 a
Molecular mapping of deletion breakpoints by Southern blotting Fig. 13.7 b, c
Duplications add material to the genome Fig. 13.8 a,b
Duplication loops form when chromosomes pair in duplication heterozygotes • In prophase I, the duplication loop can assume different configurations that maximize the pairing of related regions Fig. 13.8 c
Duplications can affect phenotype • Novel phenotypes • More gene copies • Genes next to duplication displaced to new environment altering expression Fig. 13.9
Unequal crossing over between duplications increases or decreases gene copy number Fig. 13.10
The effects of duplications and deletions on phenotpye • Heterozygosity creates imbalance in gene product altering phenotypes (some lethal) • Genes may be placed in new location that modify expression • Deletions and duplications drive evolution of the genome





















