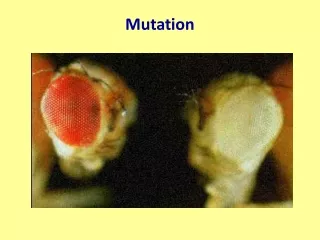
Mutation
E N D
Presentation Transcript
Mutation Dan Graur
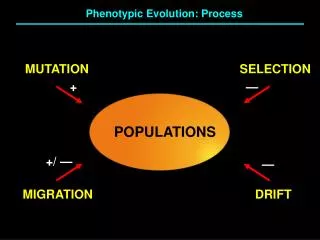
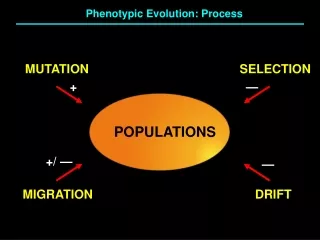
From an evolutionary viewpoint, a mutation is a DNA sequence in the germline that differs from its counterparts in somatic cells, and which is passed on from parents to progeny, thereby endowing an individual in the next generation with a novel genetic constitution. Mutations are the ultimate source of variation and novelty in evolution.
The Mutational “Balancing Act” Too many mutations No Life Too few mutations No Evolution
Mutations can occur in either somatic or germline cells. Somatic mutations are not inherited in animals. They are, therefore, evolutionarily irrelevant.
Vascular plants do not have a sequestered germline and, therefore, the distinction between somatic and germline mutations is not absolute.
Replication-dependent mutations • Replication-independent mutations
DNA sequences are usually copied exactly during DNA replication. Rarely, however, errors occur, giving rise to novel sequences. These errors are called replication-dependent mutations.
Mutations may be divided into induced and spontaneous mutations according to whether or not the mutation was caused by an external mutagen, such as caffeine, nitrous acid, ethidium bromide, or ionizing and ultraviolet radiation.
Examples of mutagens: Hiroshima Chernobyl
Mutations affect the genotype. Mutations may or may not affect the phenotype. A strict unambiguous correspondence between genotype and phenotype does not exist.
Mutations that affect the phenotype may be: amorphic (loss of function), hypomorphic (partial loss of function), hypermorphic (increase in function), neomorphic (gain of function)…
An amorphic mutation in tyrosinase and its consequences. MLLAVLYCLLWSFQTSAGHFPRACVSSKNLMEKECCPPWSGDRSPCGQLSGRGSCQNILL SNAPLGPQFPFTGVDDRESWPSVFYNRTCQCSGNFMGFNCGNCKFGFWGPNCTERRLLVR RNIFDLSAPEKDKFFAYLTLAKHTISSDYVIPIGTYGQMKNGSTPMFNDINIYDLFVWMH YYVSMDALLGGSEIWRDIDFAHEAPAFLPWHRLFLLRWEQEIQKLTGDENFTIPYWDWRD AEKCDICTDEYMGGQHPTNPNLLSPASFFSSWQIVCSRLEEYNSHQSLCNGTPEGPLRRN PGNHDKSRTPRLPSSADVEFCLSLTQYESGSMDKAANFSFRNTLEGFASPLTGIADASQS SMHNALHIYMNGTMSQVQGSANDPIFLLHHAFVDSIFEQWLRRHRPLQEVYPEANAPIGH NRESYMVPFIPLYRNGDFFISSKDLGYDYSYLQDSDPDSFQDYIKSYLEQASRIWSWLLG AAMVGAVLTALLAGLVSLLCRHKRKQLPEEKQPLLMEKEDYHSLYQSHL Pro81Leu NAIROBI, Kenya — The belief that albino body parts have magical powers has driven thousands of Africa's albinos into hiding, fearful of losing their lives to dealers who can sell their limbs for as much as $75,000.
A neomorphic mutation in a nongenic region between the a-globin genes and their upstream regulatory elements causes a-thalassemia Acquisition of a new transcription factor binding site Thesequence 5'-TAATAA-3’ changed into 5'-TGATAA-3’,creating a new binding site.
Point Substitution Deletion Mutations Insertion Substantive Segmental Inversion Contextual Recombination
transition substitutions transversion recombination deletion insertion inversion
synonymous nonsynonymous nonsense
Codons that can mutate to a termination codon by a single nucleotide substitution, e.g., UGC (Tyr), are called pretermination codons.
Mutations in stop codons causing the translation to continue are called “sense” mutations.
Each sense codon can mutate to nine other codons by means of a single nucleotide substitution. Example: CCU (Pro) can experience six nonsynonymous substitutions, to UCU (Ser), ACU (Thr), GCU (Ala), CUU (Leu), CAU (His), or CGU (Arg), and three synonymous substitutions, to CCC, CCA, or CCG.
Since the standard genetic code consists of 61 sense codons, there are: 61 9 = 549 possible nucleotide substitutions.
non- reciprocal
Reciprocal recombination is a powerful generator of variability. For example, recombinations between 5’—AACT—3’ and 5’—CTTG—3’ may result in 6 novel sequences: 5’—ATTG—3’ 5’—CACT—3’ 5’—AATG—3’ 5’—CTCT—3’ 5’—AACG—3’ 5’—CTTT—3’
The more variants there are, the more variants will come into being through recombination, and the rate of generating new genetic variation will become quite high. “Variation begets variation.” Golding and Strobeck (1983)
Insertion Insertion Unequal Crossing Over
Intrastrand Deletion
When two sequences are compared, it is impossible to tell whether a deletion has occurred in one or an insertion has occurred in the other. INsertions and DELetions are collectively referred to as INDELs.
The number of nucleotides in an indel range from one or a few nucleotides to contiguous stretches of thousands of nucleotides.
Indel lengths exhibit a bimodal frequency distribution, with short indels (up to 20–30 nucleotides) being caused by errors of DNA replication, such as slipped-strand mispairing, and with long indels occurring mainly because of unequal crossing-over, site-specific recombination, DNA transposition, or horizontal gene transfer.
Inversion (a) chromosome breakage and rejoining (b) intrachromosomal crossing-over between two homologous segments that are oriented in opposite directions
Spatial Distribution of Mutations
Mutations do not occur randomly throughout the genome. Some regions are hotspots of mutation. One such hotspot is the dinucleotide 5'CG3' (CpG), in which the cytosine is frequently methylated in many animal genomes, and may mutate to 5'TG3'.
Mutations do not occur randomly throughout the genome. Some regions are hotspots of mutation. The dinucleotide 5'TT3' is a hotspot of mutation in many prokaryotes, but not in eukaryotes.
A mutation is expected to occur with the same frequency under conditions in which it confers an advantage on the organism carrying it, as under conditions in which it confers no advantage or is deleterious. “It may seem a deplorable imperfection of nature that mutability is not restricted to changes that enhance the adeptness of their carriers.” Theodosius Dobzhansky (1970)