Download

1 / 1
30 likes | 312 Vues
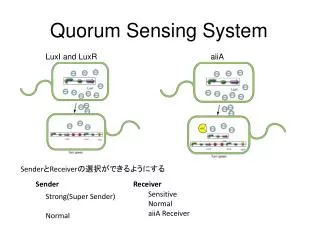
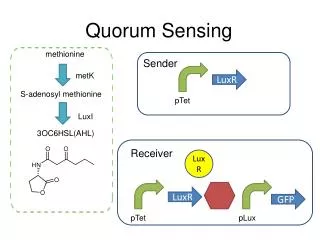
Do the two AHL inputs contribute equal noise?. 0.15. 6. Different AHL conditions with same mean response . 10. 0.1. 1 M 3OC6 HSL. Noise and Crosstalk in the Two Quorum-Sensing Channels of Vibrio fischeri. C8 = 0, 3OC6 = 200 nM /mean = 0.37. 1000. 5. 10. 0.05. 200 nM. 0. 4.

E N D
Do the two AHL inputs contribute equal noise? 0.15 6 Different AHL conditions with same mean response 10 0.1 1 M 3OC6 HSL Noise and Crosstalk in the Two Quorum-Sensing Channels of Vibrio fischeri C8 = 0, 3OC6 = 200 nM /mean = 0.37 1000 5 10 0.05 200 nM 0 4 Luminescence (at OD = 0.28/cm) 10 100 C8 HSL (nM) • P.D. Pérez, J.T. Weiss, S.J. Hagen* • Physics Department, University of Florida, Gainesville FL 32611-8440 • *Email: sjhagen@ufl.edu 3 20 nM 10 • Program # 909-Pos Poster B709 • Email: sjhagen@ufl.edu • If your phone has a QR barcode reader application you can download the abstract and author information by scanning this barcode. 0.15 C8 = 20 nM, 3OC6 = 500 nM /mean = 0.43 no 3OC6 HSL 2 0.1 10 Fraction of population 1 nM 10 100 1000 1 nM 10 100 1000 10 0.05 [C8 HSL] (nM) [3OC6 HSL] (nM) 0 0 C8 = 100 nM, 3OC6 = 800 nM /mean = 0.35 300 600 900 3OC6 HSL (nM) 0.15 no C8 HSL 10 nM C8 HSL 0.1 50 nM 500 nM 0 1000 2000 3000 4000 5000 6000 0.05 GFP expression per cell 0 BELOW: C8 HSL suppresses both mean and variance in lux response. Non-induced cells were placed in the flow chambers and supplied (starting at time t = 0) to fresh growth medium containing 3OC6 HSL + C8 HSL. All chambers contained 100 nM 3OC6 HSL. The C8 HSL concentration was 0, 500, or 1000 nM. • 1. The LuxI/LuxR system: • The marine bacterium Vibrio fischeri generates bioluminescence (light emission) through a bacterial luciferase and related enzymes that are encoded by the lux operon, luxICDABEG. V.fischeri regulates its bioluminescence through a quorum sensing (QS) mechanism, which is summarized for simplicity in the figure at RIGHT: The protein LuxI synthesizes a signal molecule (the autoinducer 3OC6 HSL – red symbol) that diffuses into the environment. When the local 3OC6 HSLconcentration reaches a high level (roughly 10-100 nM), it activates the receptor LuxR, a transcriptional activator for the lux genes. Hence, the bioluminescent output is reduced at low population densities (low 3OC6 HSL concentrations) but luminescence switches on when population density increases and 3OC6 HSL accumulates. • Abstract: • Bacteria use the chemical signaling mechanism known as quorum sensing (QS) to probe their environment. By releasing a diffusible molecule (an autoinducer) into their environment, they can use the local concentration of this molecule as an indicator of their own population density as well as other parameters, and regulate phenotype accordingly. QS regulatory pathways are often complex, receiving inputs from several different autoinducer species. It is not well understood how and why bacteria integrate QS signals from multiple channels and how the signal-to-noise property of one autoinducer channel affects that of other channels. • We have studied the role of noise and crosstalk between two QS signals in Vibrio fischeri, a bioluminescent bacterium that colonizes the light organ of some fish and squid species. V.fischeri regulates its bioluminescence with three QS signals, including a 3-oxo-C6 homoserine lactone autoinducer (3OC6 HSL) and a C8 homoserine lactone autoinducer (C8 HSL) that is also critical to colonization behavior. When both C8 HSL and 3OC6 HSL signals are present, the response of the luminescence operon is complex. Here we consider how crosstalk from the C8 HSL signal affects the response to 3OC6 HSL. We present a simple competition model that quantitatively describes the average (bulk culture) response of luxto combinations of C8 HSL and 3OC6 HSL signals. We also use microfluidic flow chambers to characterize the noise or heterogeneity in the individual cell response to the two autoinducer signals. We find that, although the robust lux response to 3OC6 HSL activation is highly heterogeneous (noisy) across a population of cells, the presence of C8 HSL quenches that response largely by switching off the brightest cells, suppressing heterogeneity. However, the noise distributions indicate that the QS circuit does not distinguish between different combinations of HSL inputs that produce the same average lux output. Therefore C8 HSL may benefit the population during colonization primarily by quenching early noisy bioluminescence response, which is poorly controlled by the 3OC6 HSL signal channel alone. Brightness percentiles (integral of GFP expression histograms) GFP expression level in individual cells (no C8 HSL) +500 nM C8 HSL +1 M C8 HSL (no C8 HSL) +500 nM C8 HSL +1 M C8 HSL t = 0.73 hrs t = 0.73 hrs t = 1.4 hrs t = 1.4 hrs t = 2.5 hrs t = 2.5 hrs Percentile 4 3.5 Fraction of Population t = 3.4 hrs t = 3.4 hrs 3 2.5 Luminescence (arb units) • 2. QS is noisy • At LEFT a group of individual Vibrio fischeri cellsare adhered in a flow chamber that supplies a fixed concentration of 1 M 3OC6 HSL autoinducer in fresh growth medium. The cells are imaged (A) in darkfield and (B) in their bioluminescent emission. The bioluminescent emission is very weak, with no more than a few hundred photons /minute/cell detected. • We find that the bioluminescence response is highly variable between cells [Pérez 2010]. For any given concentration of 3OC6 HSL in the flow chamber, individual cells differ widely in both the intensity of their luminescence and in the kinetics of their response. Even saturating (1 M) concentrations of 3OC6 HSL produce a heterogeneous luminescence response. Individual cells differ widely in the onset time (t1/2) for their luminescence (LEFT, D) and in their resulting brightness (LEFT, C). • Therefore, while the LuxI/LuxR QS mechanism provides good control over the average response, it exerts only weak control over individual cell behavior. 2 t = 3.8 hrs t = 3.8 hrs 1.5 1 t = 4.5 hrs 0.5 t = 4.5 hrs 0 0 1000 500 800 600 GFP Expression (cts/pixel/cell) GFP Expression (cts/pixel/cell) 400 C8 HSL (nM) 200 1000 0 3OC6 HSL (nM) 5. How does C8 HSL affect heterogeneity? In the experiment ABOVE the three AHL conditions cross the contours of constant lux activation. At LEFT the three experimental conditions roughly follow the contours. ABOVE: The presence of C8 HSL suppresses both the mean and the variance in GFP expression. Without C8 HSL the cells develop much broader (& non-gaussian) distribution in expression levels over time. LEFT: If conditions are chosen so that the mean expression is constant, then all three populations show very similar variance. That is, the probability distribution for expression is insensitive to the relative levels of C8 HSL and 3OC6 HSL input signals. The response does not distinguish between signal combinations such as (blue) low 3OC6 HSL + low C8 HSL versus(red) high 3OC6 HSL + high C8 HSL. C 4. A simple competition model We can model (ABOVE LEFT) the crosstalk between the C8 HSL and 3OC6 HSL signals. LuxR forms a complex (actually a multimer) with either C8 HSL or 3OC6 HSL. However, only the LuxR-3OC6 HSL complex is a strong transcriptional activator for lux. C8 HSL competes for LuxR and quenches lux response at high concentrations of 3OC6 HSL. Hence, unlike V.harveyi luminescence, where two QS signals equally and symmetrically activate lux [Mehta 2009], these two signals in V.fischeri act in opposition. The experimental data (ABOVE RIGHT) can be fit to the model to give the dissociation constants (roughly Keq10-30 nM for both LuxR-3OC6 and LuxR-C8 HSL). The two AHL inputs contribute differently to lux activation. How does the C8 HSL input affect noise and heterogeneity in lux? The contour plot (LEFT) shows the luminescence output as a function of 3OC6 HSL and C8 levels. We measured lux activity in individual cells in order to see how the noise in lux varies as we cross (white circles, LEFT) or follow (gray circles) the contours. For this experiment we do not measure single-cell bioluminescence. Rather we measure the fluorescence of a gfp reporter inserted into the lux operon (strain JB10). As a reporter of lux activation in individual cells, GFP is orders of magnitude brighter than bioluminescence. D • 3. V.fischeridetects multiple QS signals • A more complete description of QS regulation of V.fischeri bioluminescence includes three distinct autoinducer signals, represented by the red symbols in the diagram ABOVE. The homoserine lactones 3OC6-HSL and C8-HSL exert the greatest regulatory effect on the bioluminescence genes (lux operon). 3OC6 HSL and C8 HSL are synthesized by LuxI and AinS respectively and are detected by the receptors AinR andLuxR. Although 3OC6 HSL acts rather directly (via LuxR) to activate the lux operon, the action of C8 HSL is less direct, as it operates via a phosphorelay cascade involving LuxU, LuxO, LitR, and an sRNA. The C8 HSL and 3OC6 HSL signals are believed to act sequentially during the bacterium’s colonization of its animal host, as C8 HSL affects colonization factors in addition to bioluminescence [Lupp et al. 2003; Lupp and Ruby 2005]. • However it is not understood why bioluminescence should require multiple signal inputs (30C6 HSL, C8 HSL, and AI2). What information does the individual cell receive from additional inputs (such as C8 HSL) and how does this extra signal interact with the bioluminescence response to 30C6 HSL? • The figure BELOW shows the bioluminescence response of a bulk culture of V.fischeri in the presence of both C8 HSL and 3OC6 HSL autoinducers. Crosstalk is clearly apparent as the behavior is complex: 3OC6 HSL switches bioluminescence on, and the addition of C8 HSL also has a weak activating effect at low 3OC6 levels. But at high 3OC6 levels, C8 suppresses the bioluminescence, so that the two signals act antagonistically. • (Response at t = 3.6 hr after introduction of HSL signals) • 6. ConclusionsWe have studied the question of why V.fischeri uses three autoinducer signals to regulate its bioluminescence. Although the C8 HSL signal pathway activates the expression of the LuxR receptor, the primary effect of C8 HSL on lux activation seems to be its competitive inhibition of the interaction of the 3OC6 HSL signal with LuxR. When 3OC6 HSL is present, the addition of C8 HSL effectively suppresses lux activation. We find that a simple competitive binding model provides a satisfactory fit to the experimental data, although more sophisticated ODE models can also reproduce this behavior [Kuttler and Hense 2008]. The model illustrates the highly asymmetric action of the two signal input channels: C8 and 3OC6 HSL play antagonistic roles in controlling lux. Furthermore, our single-cell studies show that the 3OC6 HSL signal exerts poor control of the lux response, which is extremely noisy (heterogeneous) across a population of cells. The action of C8 is to quench lux activation by collapsing this heterogeneous distribution. Therefore one biological benefit of C8 HSL regulation of bioluminescence may be to dampen anomalously bright ("rogue") cells early in colonization, when emission is unneeded. However the QS system does not seem to distinguish between different chemical combinations of C8 HSL and 3OC6 HSL inputs if these correspond to the same average output: different signal combinations that generate the same average output also generate the same probability distribution of luxactivation levels. Microfluidic Flow chamber: Cells are observed in 3 parallel flow chambers that supply different combinations of AHL signals • Materials and methods • Strains: For our studies of noise in LuxI/LuxR bioluminescence, we used a wild-type Vibrio fischeri strain MJ11. The individual cell bioluminescence is a weak signal however and difficult to measure with precision on short (< 10-15 minutes) time scales and at low AHL levels. For our microfluidic studies using 3OC6 HSL + C8 HSL we therefore used a V.fischeri strain containing a gfp reporter for the lux operon: Vibrio fischeri strain JB10 is an ES114-derived mutant containing a gfp insertion (luxI-gfp-luxCDABEG) that provides a fluorescent GFP readout when activated [Bose 2007]. • Microfluidics: The microfluidic device consists of three parallel flow chambers, each of which supplies a constant flow of (defined) growth medium containing a fixed concentration of exogenous 3OC6 HSL and/or C8 HSL. AHLs produced by the cells in the chamber are washed away rapidly by the flow and do not accumulate in the device. The microfluidic devices are fabricated from polydimethylsiloxane (PDMS) by casting on a silicon wafer, and sealed with a glass coverslip. Each flow chamber in the device is 400 m wide and typically 10-20 m deep. • Observation of cells: Cells in exponential phase but low population density (i.e. not autoinduced) are injected into the device and adhere to a coating of poly-L-lysine on the coverslip (= the window of the device). The flow chamber is then placed on the stage of a Nikon TE2000U inverted microscope at 22 C and observed at 20x or 30x in both phase contrast and GFP fluorescence. Images are recorded with a Princeton Instruments Coolsnap HQ2 camera. We use Matlab to identify and select individual cells from the phase contrast images and then calculate the brightness of those cells in the corresponding fluorescence images. Flow to chamber #1 Chamber length Chamber width 10 mm 0.40 mm Flow to chamber #2 Flow to chamber #3 • Acknowledgment • The authors thank Prof. Mark Mandel for providing strain MJ11 (wild type) and Prof. Eric Stabb for providing mutant JB10 (+gfp reporter) of V.fischeri. Funding support through NSF MCB 0347124 is gratefully acknowledged. • References • Bioluminescence in Vibrio fischeri is controlled by the redox-responsive regulator ArcA, J.L. Bose et al., • Molec. Microb. (2007) 65 538–553. • Interplay of two QS regulation systems of V.fischeri, C.Kuttler & B.Hense, J. Theor. Biol. (2008) 251 167-180. • The Vibrio fischeri QS systems ain and lux sequentially induce luminescence gene expression and are • important for persistence in the squid host, C. Lupp et al., Molec. Microb. (2003) 50 319-331. • Vibrio fischeri uses two quorum sensing systems for the regulation of early and late colonization • factors, C. Lupp and E.G. Ruby, J. Bact. (2005) 187 3620-3629. • Information processing and signal integration in bacterial quorum sensing, P.Mehta, S. Goyal, T. Long, • B. L. Bassler, and N.S. Wingreen, Mol. Syst. Biol. (2009) 5 1-11. • Heterogeneous response to a quorum-sensing signal in the luminescence of individual Vibrio fischeri, • P. Pérez and S.J. Hagen. PLoS ONE (2010)5e15473. Lux response to two QS signals: C8 HSL and 3OC6 HSL V. fischeri strain JB10 in flow chamber, viewed in (LEFT) GFP fluorescence and (RIGHT) Phase Contrast