Download

1 / 38
380 likes | 622 Vues
Single-molecule DNA Manipulations. Course Overview. Day 1: Techniques & Basic Results Day 2: Twisting DNA molecules Day 3: DNA and RNA polymerases Day 4: DNA topoisomerases Day 5: DNA packaging. Day 1: Techniques & Basic Results. Historic overview & Introduction

E N D
Course Overview • Day 1: Techniques & Basic Results • Day 2: Twisting DNA molecules • Day 3: DNA and RNA polymerases • Day 4: DNA topoisomerases • Day 5: DNA packaging
Day 1: Techniques & Basic Results • Historic overview & Introduction • Measurement techniques, calibration, noise • Stretching nucleic acids • Single-molecule DNA sequencing? • Zero-force experiments (fluorescence measurements)
Bacteria ~eV ~kBT (4 x 10-21 J) or 4 pN nm (1.6 x 10-19 J) Length, energy- and force-scales Energies
Rotation of the F1-ATPase Noji et al., Nature (1997) 386: 299-302.
Object localized in space Real-time readout “Synchronized” Reversible Object freely diffusing Offline readout “Unsynchronized” Irreversible Single-molecule paradigm Single-molecule assay Bulk biochemical assay
Fixed-position Atomic Force Microscope Micropipette Optical Tweezer Fixed-force Magnetic Trap Instruments Feedback loop can convert one into the other
Atomic Force Microscope Cantilevers: ~50-100 mm long, 30 mm wide, 0.2 mm thick High spatial positionning accuracy (0.1 nm) Very stiff (5-100 pN/nm) cantilever High-force instrument (~10 pN-1 nN) High bandwidth (~kHz in water) Large size and high bandwidth lead to large noise: 5-10 pN rms noise with 1 kHz bandwidth High noise is due to size ( ~0.5 pN/Hz-1/2) smaller cantilevers being developped • Measurement modes (imaging, stretching)
Micropipette Weaker cantilever (~2 pN/mm) Larger cantilever, lower bandwidth lower force noise than AFM High forces achievable Rotation also possible (Bustamante, Heslot)
Optical Trap Relatively stiff (~0.1-1 pN/nm) High Forces (~100 pN) Small beads (~0.5 mm) High bandwidth (~kHz) Low force noise Rotation possible but difficult Bead trapped at beam waist Higher dielectric than water needed
Magnetic Tweezer Very weak stiffness of “trap” (pN/mm) results in a “constant force” mode requires a stiff tether Force depends on bead size: 1 mm dia bead ~ 1 pN 2.8 mm dia bead ~ 15 pN 4.5 mm dia bead ~ 80 pN Bandwidth depends on bead size Low force noise, ultra stable and very low drift Rotation easy
Other techniques • Flow fields • Force changes along DNA • Costly in protein • Electric fields • Force changes along DNA.
Force Calibration • Calibration against flow field (F=6phrv) • Micropipettes/AFM cantilevers can be calibrated using a set of levers of decreasing stiffness • Trap stiffness in all cases easily determined by analyzing Brownian motion
Fx= Fsinq ~ Fq ~ F dx __ l F F __ __ l l 1 1 _ _ <dx2> = kBT kx<dx2> = kBT 2 2 Brownian Motion Analysis (Tethered bead in a harmonic potential) _ Fx = dx = kxdx Equipartition of energy: kBT l ____ F = <dx2>
Signal-to-noise Thermal agitation causes the mean force to fluctuate with a variance <dF2> = 4kBT 6phr Df For a ~1 mm diameter bead in water (h = 10-3 poise) at room temperature, dF ~ 10 fN/Hz1/2 The detector (bead, cantilever) undergoes rms fluctuations in its mean position: dz = dF/kz (for DNA in our experiments k ~ 10-8 to 10-7 N/m) dz ~ tens of nm with ~1 s averaging • To reduce noise, three approaches (each with its own problems): • Average longer • Smaller detector • Stiffer (i.e. shorter) DNA or polymer
l 3 _ k BT ___ _ 2 x l0 Polymer Springs • Polymer elasticity characterized by • Persistence length x = A/kBT • Relative extension l/l0 • Simplest case: random walk (Freely-Jointed Chain, FJC) • Fully flexible joints between persistence-length units (no bending energy) • Entropic elasticity at low force: F= • High force is like aligning spin with mag.field
Freely-Jointed Chain vs. Worm-like chain A: entropic B: enthalpic C: “overstretch” 1st measurement: Finzi et al., Science (1992) 258:1122-6.
Effect of ionic conditions on x Fit is to Poisson-Boltzmann model for uniformly charged cylinder Baumann et al., PNAS (1997) 94:6185-90.
BS transition and ssDNA C B D A Cluzel et al., Science (1996) 271:792-4. Cui et al., “ “ “ 795-9.
Protein elasticity:unfolding titin Rief et al., Science (1997) 276 :1109-12.
Stretching ssDNA Dessinges et al., PRL (2002) 89, 248102.
Base-pairing disrupted by salt or chemical modification • = 0.8 nm • Y ~ 200 Mpa • Electrostatics+pairing • Self-avoiding ssDNA FJC: Dessinges et al., PRL (2002) 89, 248102.
Single-molecule sequencing? • l-Exonuclease • a-hemolysin/nanopores (D. Branton Harvard/Rowland) • Optical waveguides (W. Webb, Cornell U) Human genome: ~3 Gbp…how to sequence in one hour??
Digestion of DNA by l-Exonuclease “Direct” assay “Conversion” assay Perkins et al., Science (2003) 301: 1914-8. Van Oijen et al., Science (2003) 301:1235-8.
Constant-force digestion rate Perkins et al., Science (2003) 301: 1914-8.
Digestion rate is force-independent Perkins et al., Science (2003) 301: 1914-8.
Conversion of dsDNA to ssDNA Van Oijen et al., Science (2003) 301:1235-8.
Waveguide Sequencing Levene et al., Science (2003) 299: 682-6.
Folding/unfolding of structured RNA Liphardt et al., Science (2001) 292: 733-7.
Folding/unfolding of structured RNA DG ~ F1/2Dx = 14pN x 20 nm = 280 pNnm = 70 kBT (or, ~170 kJ/mol) Liphardt et al., Science (2001) 292: 733-7.
Effect of an external potential on rates Energy landscape (no external force) Energy landscape (including external force) Energy Dln Dld Potential energy from external force Reaction coordinate (distance, l) along stretching force a(F) = a0 exp(FDln/kBT) b(F) = b0 exp(-FDld/kBT)
Unzipping DNA Essevaz-Roulet et al. PNAS (1997) 94 11935-11940 Spatial resolution ~ 50 nm Needle stiffness ~ 1.7 pN/mm If Eu ~ 2kBT and Dxu ~ 2x0.3nm, we expect Fu ~ 13 pN
Unzipping signal and force flips Essevaz-Roulet et al. PNAS (1997) 94 11935-11940
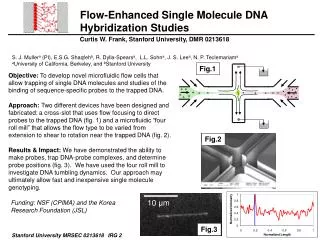
TIRF-based detection of DNA hybridization Singh-Zocchi et al., PNAS (2003) 100: 7605-10.
Hybridization signal Singh-Zocchi et al., PNAS (2003) 100: 7605-10.
Combining manipulation and visualization: actin/myosin interaction Ishijima et al. Cell (1998) 92:161-71.
Combining manipulation and visualization: DNA unzipping Lang et al., J Biol. (2003) 2 :6