Single molecule FRET
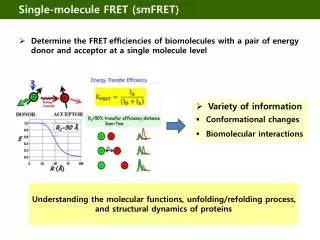
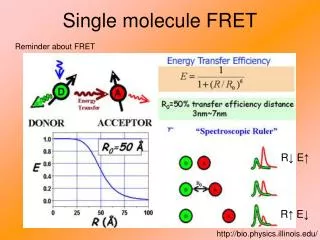
Single molecule FRET. Reminder about FRET. R↓ E↑. R↑ E↓. http://bio.physics.illinois.edu/. smFRET on Telomeric DNA Folded [G-quadruplex: 6- telomeric sequence (TTAGGG) 4 ] Unfolded. G-quadruplex: at least 3G, repeated 4 times. Telomeres: single-stranded ends of DNA.

Single molecule FRET
E N D
Presentation Transcript
Single molecule FRET Reminder about FRET R↓ E↑ R↑ E↓ http://bio.physics.illinois.edu/
smFRET on Telomeric DNAFolded[G-quadruplex: 6- telomeric sequence (TTAGGG)4] Unfolded G-quadruplex: at least 3G, repeated 4 times Telomeres: single-stranded ends of DNA. Fold up to make sure not recognized as double-stranded breaks. G-quadruplex “hides” the double-stranded break of DNA
G-Quadruplex DNA Salt stabilizes G-quadruplex Excite 1 wavelength, look at D-only and A-only. At given [salt] will have stochastic (random) amount of Energy Transfer
Lots of approximations In fact ID= ID/qD
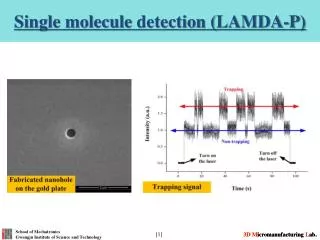
Imaging (Single Molecules) with very good S/N (at the cost of seeing only a thin section very near the surface) TIR- (q > qc) Exponential decay Total Internal Reflection (TIR) Microscopy dp=(l/4p)[n12sin2i) - n22]-1/2 For glass (n=1.5), water (n=1.33): TIR angle = >57° Penetration depth = dp = 58 nm With dp = 58 nm , can excite sample and not much background.
DNA Sample Cy-3 Dye Biotinylated DNA 3’ Streptavidin Biotinylated BSA coverslip coverslip Experimental Setup: Imaging Single Molecules Total Internal Reflection Microscopy Cy-5 Dye
Sample Objective (NA=1.49) Dichroic Laser Filter Lens CCD Detector Experimental Setup: Imaging Single Molecules Total Internal Reflection Microscopy Prism TIR (Wide-field) Objective TIR (Wide-field) Excite donor-only (or can use two wavelength, i.e. ALEX) Objective: + somewhat higher collection efficiency, (-) autofluorescence of objective
G-Quadruplex DNA Excite 1 wavelength, look at D-only and A-only. At given [salt] will have stochastic (random) amount of Energy Transfer
HMM Analysis Determine the average transition state between two conformations. Assume Markov state where probability per unit time depends only on current state the lifetime of each state is exponential. Process is hidden because we don’t observe FRET state, but rather the fluorescence
What is Hidden Markov Method (HMM)? Hidden Markov Methods (HMM) –powerful statistical data analysis methods initially developed for single ion channel recordings – but recently extended to FRET on DNA, to analyze motor protein steps sizes – particularly in noisy traces. What is a Markov method? the transition rates between the states are independent of time. Why is it called Hidden? Often times states have the same current, and hence are hidden. Also, can be “lost” in noise. What is it good for? Can derive signals where it appears to be only noise! Lec 17: 4/2010, Physics 498: http://courses.physics.illinois.edu/biop401/lecturesSP10.htm
C O → ← Simple model (non-HMM) applied to ion channels Transitions between one or more closed states to one or more open states. (From Venkataramanan et al, IEEE Trans., 1998 Part 1.) Model (middle) of a single closed (C) and open (O) state, leading to 2 pA or 0 pA of current (middle, top), and a histogram analysis of open (left) and closed (right) lifetimes, with single exponential lifetimes. In both cases, a single exponential indicates that there is only one open and one closed state. Hence the simple model C O is sufficient to describe this particular ion channel. In general, N exponentials indicate N open (or closed) states. Hence the number of open (closed) states can be determined, even if they have the same conductance. In addition, the relative free energies of the open vs. closed two states can be determined because the equilibrium constant is just the ratio of open to closed times and equals exp(-DG/kT).
Outline con’t: Making a Markov Process Two general points: Notice that correlations between states are inherent in the topology and rate constants. (For example, if state B can only be reached through State A, then the presence of state B would indicate that state A should be found in some time before State B.) Furthermore, like the histogram method mentioned, multiple states (conformations) with the same signal (e.g. multiple open states with the same ionic conductance, or multiple closed states) can be detected via statistical analysis from their lifetime distributions.
c. a. b. Hidden Markov Models An impressive feature of combining Hidden Markov & Maximum Likelihood models is the ability to extract signal from noise. Indeed, they are often called Hidden Markov Methods because the observable (ionic current, or position in the case of molecular motors) is often hidden in the noise. Use of Hidden Markov Methods to analyze single ion channel recordings. a) Ideal current vs. time, showing ion channel transitions with two different conductivities and forward and backward rate constants of 0.3 and 0.1. B) Data of (a) added to white noise such that noise level = signal level. C) Extraction of kinetic parameters using HMM from noisy data in b, showing kinetic constants can be recovered. (Venkataramanan et al., IEEE Transactions, 1998, Part I.)
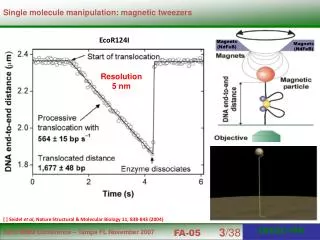
Molecular motors can be modeled like ion channels Conceptually a transition between a closed and open state in an ion channel corresponds to a molecular motor taking a step. A transition between one closed ion channel state and another closed state corresponds to a conformational change within the motor that does not lead to a step – e.g. hydrolysis of ATP into ADP + Pi in kinesin. These states can potentially be detected because they affect the kinetics (stepping rate and /or distribution). Hence, an important aspect of this analysis is that we may be able to discover the presence of new conformational states, likely associated with nucleotide states. In addition, if motors walk in a hand-over-hand mechanism, and there is an asymmetry between the two heads – e.g. a step where “left foot” goes forward, vs. “right foot” goes forward – and one tends to overwinds the coiled-coiled stalk, and the other tends to underwind the stalk, then this may show up in the stepping kinetics. This is analogous to an ion channel with multiple states of equal conductance.
Two kinesins operate simultaneously in vivo Must use Hidden Markov Method to see Syed, unpublished Unlikely due to microtubule motion because fairly sharply spiked around ±4-5 nm Two kinesins (+2 Dyneins), in vivo, are moving melanosome