Membrane Structure and Function in Cells
Explore the intricacies of lipid bilayers, membrane proteins, and cell membrane composition. Learn about the importance of membrane asymmetry, phospholipids, cholesterol, and cellular signaling.


Membrane Structure and Function in Cells
E N D
Presentation Transcript
Membrane Structure ECB Chapter 11
The lipid bilayer • Membrane proteins
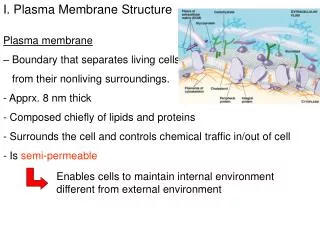
11_01_Cell.membranes.jpg Simplest bacteria Eucaryotic cells Plasma membrane: a highly selective barrier. Without membranes there would be no cells, thus no life.
11_02_plasma.membra.jpg The plasma membrane is involved incell signaling, thetransportof small molecules, andcellular growthand motility
The internal membranes 11_03_compartments.jpg The internal membranes forms the many different compartments (organells) in a eucaryotic cell Nucleus and mitochondria: enclosed by two membranes
The internal membranes • forms the many different compartments (organelles) in a eucaryotic cell • act as more than just barriers: subtle differences among them, especially differences in the resident membrane proteins, give each organelle its distinctive character
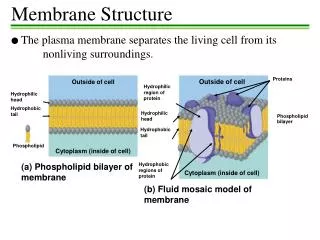
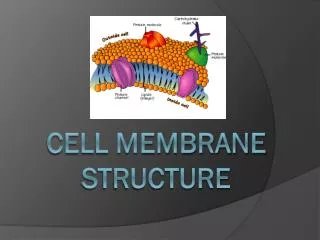
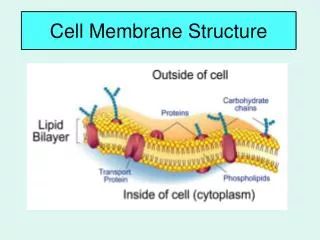
11_04_membrane.view.jpg human RBC (cross section, TEM) All cell membranes are composed of lipids and proteins and share a common general structure.
11_05_tails.philic.pho.jpg • amphipathic: • molecules with both hydrophilic and hydrophobic properties (one or two) hydrocarbon tails • A typical membrane lipid molecule has a hydrophilic head and hydrophobic tails. • The most abundant lipids in cell membranes are the phospholipids, the most common type of phospholipid in most cell membranes is phosphatidylcholine.
phosphatidylcholine: the most common type of phospholipid in most cell membranes 11_06_Phosphatidylch.jpg originate as fatty acids (hydrocarbon chains with a –COOH group at one end) (space-filling model) “phosphatidy-”: phosphate-glycerol-fatty acid
Different types of membrane lipids— all amphipathic 11_07_amphipathic.jpg serine phosphate glycerol
11_08_hydrophilic.jpg (polar molecule) (polar molecule) hydrophilic:charged atoms form electrostatic bonds or hydrogen bond with water molecule
11_09_hydrophobic.jpg (non-polar molecule) cagelike structure: highly ordered (polar molecule)
11_10_Fat_phospholip.jpg a phospholipid molecule, amphipathic a fat molecule, entirely hydrophobic
11_11_bilayer.in.H20.jpg (computer simulation)
liposome: synthetic lipid bilayer form by pure phospholipid 11_13_liposomes.jpg (to 1 mm) flexibility of lipid bilayer
11_14_synth.phospho.jpg ~1 mm
The lipid bilayer is a two-dimensional fluid 11_15_Phospho,move.jpg fluidity (usually with proteins to facilitate the process) (frequent and rapid) temperature ↓ movements ↓: (thermal motions)
fluidity of a cell membrane (at a given temperature) • phospholipid composition • the nature of the hydrocarbon tails • the closer and more regular the packing of the tails, the more viscous and less fluid • hydrocarbon chain length(↓ fluidity ↑) • the hydrocarbon tails of membrane phospholipid molecules vary in length between 14 and 24 carbon atoms, with 18-20 atoms being most usual • degree of unsaturation(double bond)(↑ fluidity ↑) • --bacterial and yeast cells-- • cholesterol(an amphipathic molecule) (in animal cells) • short, rigid molecules are present in large amounts in the plasma membrane • fill the spaces between neighboring phospholipid molecules left by the kinks in their unsaturated hydrocarbon tails • tens to stiffen the bilayer, making it more rigid and less permeable
Cell membranes are generally asymmetrical 11_17_asymmetic.dist.jpg • Phospholipids and glycolipids are distributed asymmetrically in the plasma membrane lipid bilayer. • All of the glycolipid molecules are in the external monolayer of the membrane. • Cholesterol is distributed almost equally in both monolayers. • Proteins are embedded in the bilayer with a specific orientation, which is crucial for their function.
in eucaryotic cells, new phospholipid molecules are synthesized by enzymes bound to the part of the ER membrane that faces the cytosol The lipid asymmetry is established at the point of manufacture. 11_18_Flippases.jpg selectively • one-sided insertion and selective flippases are not the only ways of producing asymmetry in lipid bilayers • glycolipids?
in eucaryotic cells nearly all new membrane synthesis occurs in ER • the new membrane assembled there is exported to the other membranes of the cell through a cycle of vesicle budding and fusion • all cell membranes have distinct “inside” and “outside” faces: the cytosolic face is adjacent to the cytosol; the noncytosolic face is exposed to either the cell exterior or the interior space of an organelle 11_19_memb.vesicles.jpg noncytosolic face cytosolic face
Lipid asymmetry is generated inside the cell • glycolipids are located mainly in the plasma membrane • they are found only in the noncytosolic half of the bilayer (expose to the exterior of the cell) • they form part of a continuous protective coat of carbohydrate that surround most animal cells • these molecules acquire their sugar groups in the golgi apparatus • the enzymes that add the sugar groups are confined to theinside of thegolgi apparatus so that the sugars are added only to lipid molecules in the noncytosolic half • no flippases to transfer the glycolipids to the cytosolic monolayer
inositol phospholipids (phosphatidylinositol) • are minor components of the plasmamembrane • play a special role in relaying signals from the cell surface to the intracellular components that respond to those signals • they act only after the signal has been transmitted across the plasma membrane • they are concentrated in the cytosolic half of this lipid bilayer
Plasma membrane proteins have a variety of functions 11_20_memb.proteins.jpg • most membrane functions are carried out by membrane proteins • in animals, proteins constitute about 50% of the mass of most plasma membrane • functions:
Functions of membrane proteins: • transport particular nutrients, metabolites, and ions across the lipid bilayer • anchor the membrane to macromolecules on either side • function as receptors that detect chemical signals in the cell’s environment and relay them to the cell’s interior • work as enzymes to catalyze specific reactions
Membrane proteins associate with the lipid bilayer in various ways 11_21_proteins.associ.jpg integral membrane proteins (directly attached to membranes) peripheral membrane proteins hydrophilic amphipathic α helix hydrophobic covalently attached lipid groups (noncovalent protein-protein interactions) hydrophilic rolled-up β sheet (β barrel)
11_22_peptide.bonds.jpg peptide bond: polar and hydrophilic
11_23_helix.cross.LB.jpg α helix: the most common form in which a polypeptide chain crosses a lipid bilayer In many transmembrane proteins the peptide chain crosses the membrane only once. Many of these proteins are receptors. about 20 a.a. are required to completely traverse a membrane in this way
A transmembrane hydrophilic pore can be formed by multiple α helices: a series of α helices that snake across the bilayer a number of times 11_24_hydrophl.pore.jpg function in the selective transport of small water-soluble molecules across membranes one or more of the transmembrane regions are formed from α helices that contain both hydrophobic and hydrophilic a. a. side chain
hydrophilic; aqueous channel 11_25_Porin.proteins.jpg • β barrel: • a β sheet curved into a cylinder, forming an open-ended keglike structure • β barrels can form only wide channels, • a β barrel is less versatile than a collection of α helices hydrophobic ex:porin protein, form large, water filled pores in mitochondrial and bacterial membranes, allow the passage of nutrients and small ions across their outer membrane while preventing the entry of large molecules such as antibiotics and toxins
detergents: • are small, amphipathic, lipidlike molecules • have both a hydrophilic and a hydrophobic region • differ from membrane phospholipid for having a single hydrophobic tail • in water, they tend to aggregate into small clusters called micells, rather than forming a bilayer as do the phospholipid 11_26_SDS_TritonX.jpg hydrophilic hydrophobic (strong ionic detergent) (mild nonionic detergent)
Membrane proteins can be solubilized with mild detergents 11_27_Triton X-100.jpg
The complete structure is known for a few membrane proteins • the standard direct method: X-ray crystallography • ex: bacteriorhodopsin photosynthetic reaction center
bacteriorhodopsin& photosynthetic reaction center • bacterial membrane proteins • with important roles in the capture and use of energy from sunlight
bacteriorhodopsin • 250 a. a. • with important rolls in capture and use of energy from sunlight • found in large amounts in the plasma membrane of an archaebacterium (Halobacterium halobium) • act as a membrane transport protein that pumping H+ out of the bacterium • gets its energy directly from sunlight for the pumping of H+
bacteriorhodopsin: act as a proton pump 11_28_Bacteriorhodop.jpg seven α helices polar a.a. side chain • generating a [H+]gradient (energy store) • driving ATP synthase (a membrane protein) generating ATP • retinal: • a light-absorbing non-protein small hydrophobic molecule • deep purple color
bacterial photosynthetic reaction center • a large complex composed of 4 protein molecules- M, L, H (transmembrane proteins) and a cytochrome(peripreral membrane protein) • taking in light energy absorbed by chlorophyll and producing high-energye- required for photosynthetic reactions
bacterial photosynthetic reaction center 11_29_PRC.jpg chlorophyll e- carrier group
The plasma membrane is reinforced by the cell cortex • A cell membrane by itself is extremely thin and fragile. • Most cell membranes are strengthened and supported by a framework of proteins, attached to the membrane via transmembrane proteins. • The shape of the cell and the mechanical properties of the plasma membrane are determined by a meshwork of fibrous proteins- the cell cortex that is attached to the cytosolic surface of the membrane.
Human RBC (SEM) 11_30_blood.cells.EM.jpg The human RBCs lack a nucleus and other intracellular organells.
The main component of cell cortex in human RBC is the protein spectrin, a long, thin, flexible rod about 100 nm in length. • In humans and mice that have genetic abnormalities in spectrin structure are anemic: they have fewer RBCs than normal, and the red cells they do have are spherical instead of flattened and are abnormally fragile.
11_31_spectrin.jpg A spectrin meshwork forms the cell cortex in human RBCs, it provides support for the plasma membrane and maintains cell shape.
Mostof proteins in the plasma membrane have sugars covalently attached to them. glycoprotein:protein links with short chain(s) of sugars(oligosaccharides, <15) proteoglycan:protein links with one or more long polysaccharide chains 11_32_sugar.coated.jpg glycoproteins, proteoglycans, and glycolipids form a sugar coating- called the carbohydrate layer
cell-surface carbohydrates function in • protection • lubrication • cell-cell recognition and adhesion • ex: lectins, egg-sperm, inflammatory responses
11_33_neutrophils.jpg Carbohydrate on the surface of WBC (neutrophils) is recognized by a lectin
Plasma membrane proteins can move within the plane of the lipid bilayer 11_34_hybrid cells.jpg
membrane domains: functionally specialized regions 11_35_lateral.mobility.jpg extrcellular matrix molecular cell cortex proteins on the surface of another cell diffusion barriers (ex: tight junction)