Download

1 / 48
480 likes | 815 Vues
Mutations and DNA Repair. Dr. Muhammad Zeeshan Hyder. Genomic Insult. DNA is heredity material of most of the organisms. Evolved due to its stability in storing information compared to RNA Many endogenous and exogenous factors insult it during the life span of each cell

E N D
Mutations and DNA Repair Dr. Muhammad Zeeshan Hyder
Genomic Insult • DNA is heredity material of most of the organisms. • Evolved due to its stability in storing information compared to RNA • Many endogenous and exogenous factors insult it during the life span of each cell • Highly active environment of cell interior (endogenous) • Environmental factors (exogenous)
Endogenous Factors causing Damage to DNA • Endogenous damage to DNA is quite high compared to exogenous. • For example in human cells at 37C about 18000 purine residues are lost in each cell due to breakage of glycosidic bond between the bases and deoxyribose ring. • Another constant turmoil at physiological pH is transformation of cytosine residues to uracil by deamination process. • About 100 to 500 times it happens in each cell per day. • In addition, oxygen free redicals which are produced in cellular metabolism frequently react with DNA and cause damage to it.
Methylating agents also damage DNA i.e. S-Adenosylmethionine (SAM), an natural methylating agent in cell, methylate DNA bases. • It methylates at adenine about 1200 times per day in each human cell. • Spontaneous deamination of adenine to hypoxanthine and cytocine to uracil. • DNA replication process also incorporates many errors in DNA
Exogenous factors causing DNA damage • Ionizing radiations from the sun can promote ring opening reactions in bases. • UV light causes fusion of adjacent pyrimidines bases to one another. That causes mutagenic and toxic lesions that must be removed or handled in some ways to avoid cell death. • Radioactive radiation from Earth can break the backbone of DNA. • There are many chemicals which can react and alter the structure of DNA and its chemistry. These are called mutagenic compounds. • Tobacco smoke contains many mutagenic compounds which induces lesions to DNA in the lungs cells.
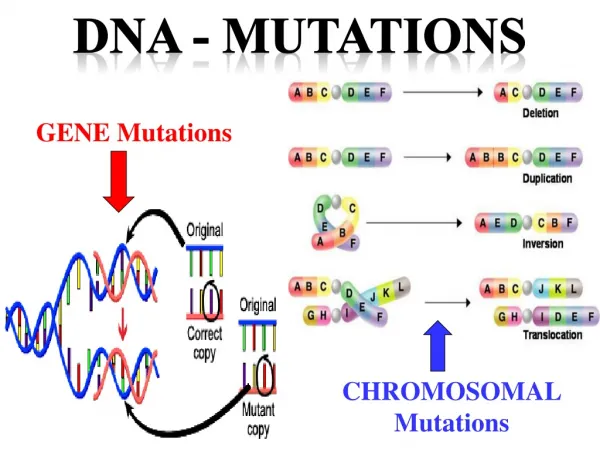
Mutations • A mutation is a heritable change in the sequence of an organism’s genome. • The mutations are of many types based upon sequence alteration they cause in genome. • The mutations may be on small scale like: • Point mutations (base substitution) • Insertion or deletion mutation • Large scale mutations may be: • Deletion and insertions of long sequence • Inversion • Duplications • In multichromosomal organisms mutations may be: • Homologous recombination • Chromosomal rearrangements • Change in the number of chromosomes.
Phenotypically mutations may be: • Null mutations (complete loss of function) • Partial-loss-of-function mutations. • Alteration-of-function mutations • Gain-of-function mutations.
Base Substitution Mutations • Point mutations that results from the substitution of one base pair for another (or for one base for another in single stranded DNA genomes) are called as base substitution mutations. • These can be further classified into: • Transition mutations (change of one purine for another or one pyrimidine to another one) or (G-A, A-G, C-T, or T-C) • Transversion mutations (interchange of a purine for a pyrimidine or interchange of one pyrimidine to purine) (G-T, G-C, A-T, A-C, T-A, T-G, C-A, C-G). • Frameshift mutations +(3n+1) (causing change in the reading of an ORF)
Point mutations results in the change of base sequence, if that sequence is a coding sequence than mutations can be: • Missense mutations (changing one amino acid with other one resultantly changing the function of a protien) . • Silent or neutral mutations (due to degeneracy of the genetic material some of the point mutations have no impact on coded proteins and thus results in no change in the proteins sequences). • Nonsense mutations (change the codon for an amino acid to one of the three stop codons: TAG (amber), TAA (ochre) or TGA (opal) leading to premature termination of a protein).
Biological Responses to DNA Damage • Given the huge spectrum of damage that the genome can suffer, the cells have evolved a multitude of mechanisms by which either damage is removed from the genome or effects of damage are mitigated. • The mechanisms by which cell respond are: • Reversal of base damage • Excision of damaged, mispaired or incorrect bases • Base Excision repair (BER) • Nucleotide excision repair (NER) • Mismatch repair (MMR) • Strand break repair • Single-strand break repair (SSBR) • Double strand base repair (DSBR) • Tolerance of base damage • Cell cycle checkpoint activation • Apoptosis
Direct Reversal of base Damage Pyrimidine Dimers are Split by Photolyase • UV radiation of 200 to 300 nm promotes the formation of a cyclobutyle ring between adjacent thymine resides on the same DNA strand known as thymine dimer. • Similarly thymine-cytosine and cytosine-cytosin dimers are also formed. • Such pyrimidine dimers locally distort DNA’s base paired structure so that it neither can transcribe nor replicated.
Direct Reversal of base Damage • A single pyrimidinedimer in E.coli genome if remains unrepaired can kill it. • These pyrimidinesdimer can be restore by a light-absorbing enzymes DNA photolyaseswhich is found in many prokaryotes and eukaryotes. • These enzymes are 55-65 kD monomers that bind to a pyrimidinedimer in DNA. • A prosthetic group in the enzyme either N5,N10-methyltetrahydrogfolate (MTHF) or 5-deazaflavin absorbs light of 300-500 nm and transfer it to pyrimidine ring in DNA and split it. • Photolyases bind DNA with high affinity but non sequence specifically. These can restore ssDNA or dsDNA
Direct Reversal of base Damage Alkyl-transferasesDealkylateAlkylated nucleotides • Some of cellular alkylating agents like N-methyl-N’-nitro-N-nitrosoguanidine (MNNG) alkylate guanine residue to make O6—methylguanine or O6-ethyleguanine. • The formation of these alkyl derivatives is highly mutagenic. • They cause incorporation of thymine instead of cytosine during cell replicaiton. • These lesions of DNA are repaired by O6-alkylguanine-DNA alkyltransferase, which directly transfer this groups onto them selves. • In E. coli Ada protein perform this function and also involve in the removal of alkyl groups from phosphate groups in DNA backbone.
Excision Repair Base Excision Repair (BER) • DNA bases are modified by reactions that occur under normal physiological conditions as well as through the action of environmental agents. • Spontaneoues hydrolysis of glycosidic bond results in detachment of nitgrogenous base from ribose ring. • Spontaneous deamination of adenine and cytosine produce hypoxanthine and uracil. • Ionization radiation can promote ring opening reactions in bases. • These type of damages are repaired by BER • Cell have a variety of DNA glycosilases which can cleave the glycosidic bond of altered nucleotide. • E.coliAlkA is base-excision enzyme • Resulting deoxyribose residue in backbone called aprinic or apyrimidinicsites are called AP sites (abasic sites)
Base Excision Repair (BER) • AP endonuclease cleave on one side of AP site • Exonuclease removes deoxyribose and several adjacent residues • DNA Pol fills the gap • DNA ligase joins the gap
Removal of Uracil • Spontaneous deamination of cytosine results in uracil formation in DNA • Change of G-C base to A-U base pairing • The uracil is excised from DNA by uracil-DNA glycosylase UDG (or uracil N-glycosilase UNG) • The rest of the process is similar to BER
Nucleotide Excision Repair • NER is found in all cells of prokaryotes and eukaryotes • It eliminate the damage to dsDNA by excising an olignucleotide containing the lesion that are characterized by the displacement of bases from their normal position. • Pyrimidinedimers • Addition of bulky substituent to a base. • Activated by helix distortion rather than recognition of any particular group. • In humans it is the major defense against cigarette smoke. • In prokaryotes its constitute of three units while in eukaryotes 16 units participate in this system.
In E. coli the NER works is an ATP dependent process. • Three proteins UvrA, UvrB and UvrC (UvrABC endonuclease) cleaves the damaged DNA at 7th nucleotide at 5’ position, and 3rd and 4th nucleotide at 3’ position. • This 11 to 12 nt long stretch of DNA is removed by an helicase i.e. UvrD protein. • The resultant gap is filled by DNA Pol I and ligated by ligase. • In prokaryotes the NER is a little complex • The damage is reconginze by (UvrA)2UvrB heterodimer, which binds dsDNA tightly but non sequence specifically. The scan the regular structure of dsDNA along the length utilizing ATP. • After recognition of damage part UvrB starts unwind 5bp around the lesion, which results in displacement of UvrA and binding of UvrC protein • The UvrB makes the nick at 3’ side of lesion while UvrC on 5’ side of the damage. • UvrD binds with these nicks and displace UvrC along with lesion containing oligonucleotide. • DNA Pol I starts filling the gap from 5’ site and displaces UvrB. • Finally the ligase seals the gap yielding refurbished DNA
Mismatch Repair • Any replication error that has eluded the editing functions of the various participating DNA Pols may still be corrected by a process called as Mismatch Repair (MMR). • E.coliPol I and III have error rates of 106-107 per base replicated • But observed rates are 109-1010 per base replicated • ???
Mismatch Repair • due to MMR • MMR can also correct insertions or deletions of up to 4 nt long due to slippage of one strand relative to others. • Defects in MMR can lead to hereditary nonpolyposis colorectal cancer (HNPCC) • Activated by distortion in DNA structure • Mismatch error need to establish parental strand or correct base from the wrong one • ???
Mismatch Repair • In E. coli, newly GATC palindromes remain hamimethylated until Dam methyltransferase has sufficient time to methylate the daughter strand. • MutSprotein binds a mismatched base pair or unpaired bases as homodimer • The MutS-DNA complex binds MutL a homodimer. • Muts-MutL complex translocate DNA along both sides and search for hamimethylated GATC palindromic sequence, by forming a loop of DNA upto 1000 bp on either sides, with the help of energy utilized by hydrolysis of ATPs.
Mismatch Repair • When hamimethylated GATC palindrome is found, the complex recruits MutH (a single stranded endonuclease) to make a nick on 5’ side of hamimethylated GATC palindrome • Then MutS-MutL complex recruits UvrDhelicase along with a exonuclease, both of which separates and degrades the nicked strand from the nick to beyond the mismatch. • If the nick is 5’ to the mismatch the exonuclease can be either RecJor exonuclease VII (5’-3’ exonucleases)
Mismatch Repair • If the nick is 3’ to the mismatch the exonuclease is exonuclease I (3’-5’exonuclease). • The resulting gap is filled by Pol III and sealed by ligase. • In eukaryotes the mechanism is more complicated with six homologs of MutS and five of MutL but homolog of MutH is only found in gram-negative bacteria, so they must have other proteins with similar functions • Similarly, eukaryotes may have other ways to determine parental strand but how it is not clear, may be by unsealed nicks in newly synthesized DNA.
Diseases Associated with defective Repair system • Defects in DNA-repair systems are expected to increase the overall frequency of mutations and, hence, the likelihood of a cancer-causing mutation • There are many genetic disease associated with defective DNA repair Xeroderma pigmentosum • Xeroderma pigmentosum, a rare human skin disease, is genetically transmitted as an autosomal recessive trait. • The skin in an affected homozygote is extremely sensitive to sunlight or ultraviolet light. The skin becomes dry, and there is a marked atrophy of the dermis. • Keratoses appear, the eyelids become scarred, and the cornea ulcerates. Skin cancer usually develops at several sites.
Many patients die before age 30 from metastases of these malignant skin tumors • In normal cell, half the pyrimidinedimers produced by ultraviolet radiation are excised in less than 24 hours. • In contrast, almost no dimers are excised in this time interval in cells derived from patients with xerodermapigmentosum Hereditary nonpolyposis colorectal cancer (HNPCC, or Lynch syndrome) • HNPCC results from defective DNA mismatch repair. HNPCC is not rare as many as 1 in 200 people will develop this form of cancer • HNPCC results due to defects in mismatch repair system, which leads to the accumulation of mutations throughout the genome. In time, genes important in controlling cell proliferation become altered, resulting in the onset of cancer.
The SOS Response • Agents that damage DNA, such as UV radiatin, alkylating agents, cross-linking agents and replication fork collapse, induces a complex system of cellular changes in E. coli known as the SOS response. • E. coli so treated cease dividing and increase their capacity to repair damaged DNA. • When E. coli cells are exposed to DNA damaging agents or agents which can inhibit the replication, a protein RecA after binding to the ssDNA (which is an indication of DNA demage), specifically mediates the proteolytic self-cleavage of LexA protein
The SOS Response • LexAprotein represses the expression of SOS response proteins • Its binds on SOS box a 20 nt long sequence present upstream of all proteins participates in SOS responst • This sequence functions as operator of SOS response proteins and LexAprotein it self. • During normal growth, LexA largely represses the expression of the SOS genes and it self too by binding on SOS box to inhibit the RNA polymerase to start transcription.
The SOS Response • When DNA damage is has sufficient to produce postreplication gaps, the damaged ssDNA binds to RecA to stimulate the cleavage of LexA and release of SOS boxes from repression. • The SOS response proteins express, including LexA, which is continually cleaved by RecA. • When the lesions have been eliminated, RecAceases simulating the self-cleavage of LexA, which now starts to repress the expression of SOS proteins. • The repair is called Translesion synthesis (TLS).
The SOS Response • The repair is filled by error-roneDNA Pol IV or V. • The DNA Pol V is also called Pol V mutasome tends to incorporates G about half as often as A opposite thymine dimmers and AP sites, with pyrimidines installed infrequently. • In TLS some time deletion and insertions also take place resulting in Frameshift mutations. • After synthesizing about 7 nt, the DNAPol V mutasome is replaced by Pol III holoenzyme, which replicate DNA about 1000 fold more fidelity.
The SOS Response • Similarly, Pol II with high fidelity also participate in TLS and express before the DNA Pol V, the role of DNA Pol II is to mediate the error-free TLS, and only if this process fails, it is replaced by DNA Pol V to carry out error-prone TLS. • The remaining nick after filling the gap is filled by DNA ligase.
The SOS Response • SOS repair is an error-prone and hence mutagenic process. • It is therefore a last resort that is only initiated about 50 minutes after SOS induction, if the DNA has not already been repaired by other mechanisms. • In E. coli, most mutations arise from the action of the SOS repair system • Under conditions of environmental stress, the SOS system functions to increase the rate of mutation so as to increase the rate at which the E. coli adapt to the new conditions.
Double-Stranded Break Repair • Double-stranded breaks (DSBs) in DNA are produced by ionizing radiation and the free redicals of oxidative metabolism. • DSBs are also normal intermediates of certain specialized cellular process such as: • Meiosis • V(D)J recombination in lymphoid cells. • DSBs if unrepaired can be lethal to cells or can cause chromosomal aberrations leading to cancer.
Double-Stranded Break Repair • So, efficient repair of DSBs is essential for cell viability and genomic integrity. • Cells have two general modes to repair DSBs: • Recombination repair • Nonhomologous end-joining (NHEJ) • NHEJ is a process, that directly regions DSBs • In NHEJ, the broken ends are aligned, trimmed/or filled in and strands are ligated.
Double-Stranded Break Repair • The core NHEJ machinery in eukaryotes includes: • the DNA end-binding protein Ku (a heterodimer of homologous 70 and 83 kD subunits, Ku70 and Ku80) • DNA ligase IV • And accessory protein Xrec4.
Double-Stranded Break Repair • Ku a abundant nuclear protein, binds to DSBs, whether blunt or with an overhang. • It is cell’s primary DSBs sensor • Ku binds to its entire length with dsDNA and encircles its central 3 bp segment forming a ring structure • The binding of Ku is not specific to any base, rather it fits snugly into major and minor grooves of DNA, and then permanently associates with this dsDNA.
Double-Stranded Break Repair • Two Ku-DNA complexes have been shown to diamerize so as to align each dsDNA strand with each other. • The Ku-DNA complexes, exposes aligned DNA, for the access of polymerases to fill in the gaps, to nucleases that trim excess nucleotides and for ligase IV in complex with Xrec4. • Nucleotide trimming, which causes mutation is an ATP dependent process. • The Ku is eventually released from the rejoined DNA by proteolytic cleavage.
Identification of Carcinogens by Aems test • Many forms of cancer are known to be caused by exposure to certain chemical agents that are therefore known as carcinogens • Approximately, 80% of the human cancer arises in this manner • Primary event in carcinogenesis is often damage to DNA • Carcinogens are consequently also likely to induce the SOS response in bacteria and thus act as indirect mutagenic agents.
Identification of Carcinogens by Aems test • There is high correlation between carcinogenesis and mutagenesis • There are presently over 60,000 man-made chemicals of commercial importance and about 1000 new ones are introduced each year. • To be sure the health of users it is important to know their carcinogenesis. • The standard animal test, exposure of rats or mice to high levels of potential/suspected agent and checking for cancer are expensive and require about 3 years to complete
Identification of Carcinogens by Aems test • So very few chemicals can be checked in this manner. • A test was devised to circumvent this problem by Bruce Ames (hence the name of the test after him), that is effective and rapid. • It is based on high correlation between carcinogenesis and mutagenesis.
Identification of Carcinogens by Aems test • Ames constructed special tester strains of Salmonella typhimurium • These strains are his- and cannot syhtheisze histidine and are unable to grow in its absence. • The cell’s envelopes that lack the lipopolysaccharide coating which renders the normal cells impermeable to many substances • The excision repair system is also inactivated in these tester strains.
Identification of Carcinogens by Ames test • The results of these changes is that any mutageneic substance can revert these tester strains to his+ phenotypes • In Ames test, about 109 bacterial cells of a mixture of tester strains are spread on a culture plate that lacks histidine. • A filter paper socked in compound under investigation is placed in the center of the plate and this plate is incubated for 2 days at 37°C.
Identification of Carcinogens by Aems test • The mutagenicity of a substance is scored as the number of such reverting colonies less the few spontaneously revertant colonies in the absence of the mutagen. • Many noncarcinogens are converted to carcinogens in the liver or in other tissues via a variety of detoxification reaction catalyzed by the cytochromes P450. • A small amount of rat liver homogenate is therefore included in the Ames test medium in an effort to approximate the effects of mammalian metabolism.
Identification of Carcinogens by Aems test • There is about 80% correspondence between the results of Ames test and animal test results about carcinogenesis of the compounds. • The dose-response curves are linear showing that there is no threshold concentration for mutagenesis. • Many compound were found carcinogenic by Ames test, which were frequently been exposed to humans.