Understanding Nucleotide Metabolism: Structure, Synthesis, and Functions
360 likes | 499 Vues
Learn about the importance, structure, and synthesis of nucleotides vital for DNA, RNA, and energy production. Explore purine nucleotide biosynthesis pathways and inhibitors, essential for cell proliferation and regulatory mechanisms.

Understanding Nucleotide Metabolism: Structure, Synthesis, and Functions
E N D
Presentation Transcript
Nucleotide Metabolism UNIT IV: Nitrogen Metabolism
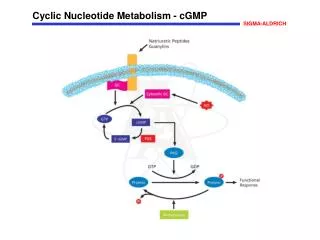
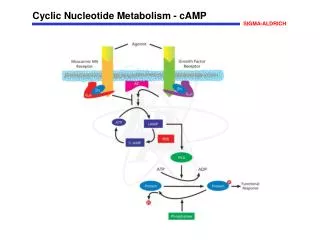
1- Overview • Nucleotides are essential for all cells • DNA and RNA synthesis/protein synthesis/cell proliferation • Carriers of activated intermediates in synthesis of some CHO’s, lipids and proteins • Structural components of several essential coenzymes, e.g., CoA, FAD, NAD+, NADP+ • cAMP and cGMP serve as second messengers in signal transduction • Energy currency • Regulatory compounds for many pathways of intermediary metabolism • Purine and pyrimidine bases can be synthesized de novo, or obtained through salvage pathways
2- Nucleotide structure • Nucleotides are composed of: • a nitrogenous base (purine/pyrimidine), • a pentose, • and 1, 2 or 3 phosphate groups. • Purine and pyrimdine structures • Both DNA and RNA contain purine bases: adenine (A) and guanine (G). • Both DNA and RNA contain cytosine (C) • DNA contains thymine (T), whereas RNA contains uracil (U). • T and U differ by one methyl group
Unusual bases are occasionally found in some species of DNA and RNA e.g., in some viral DNA, tRNA Base modifications include methylation, hydroxymethylation, glycosylation, acetylation, or reduction May aid in recognition by specific enzymes/proteins, or protect from degradation by nucleases
B. Nucleosides • Pentose sugar + N-base = nucleoside (ribonucleoside/ deoxyribonucleosides). • If sugar is ribose, ribonucleoside is produced • Ribonucleosidesof A, G, C and U = adenosine, guanosine, cytidine, uridine • If sugar is deoxyribose, a deoxyribonucleosideis produced • Deoxyribonucleosideshave the added prefix “deoxy” • eg. deoxyadenosine • Carbon and nitrogen atoms in base and sugar are numbered separately (Figure 22.3B)
Note that the carbons in the pentose are numbered 1' to 5‘ • Thus, when the 5'-carbon of a nucleoside (or nucleotide) is referred to, a carbon atom in the pentose, rather than an atom in the base, is being specified. Figure 22.3A. Pentoses found in nucleic acids.B. Examples of the numberingsystems for purine- and pyrimidine containing nucleosides.
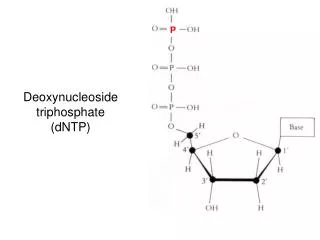
C- Nucleotides • Nucleotides are mono-, di-, or triphosphate esters of nucleosides. • The 1st phosphate group is attached by an ester linkage to the 5`-OH of pentose. • This compound is called nucleoside 5`-phosphate or 5`-nucleotide • If one phosphate is attached, the structure is a nucleoside monophosphate (NMP) e.g., AMP, CMP • If a 2nd or 3rd phosphate is added, a nucleoside diphosphate (e.g., ADP) or triphosphate (e.g., ATP) (Figure 22.4) • The 2nd and 3rd phosphates are connected by a ”high-energy” bond. • Phosphate groups give negative charges to nucleotides, and cause DNA and RNA to be referred to as “nucleic acids”.
3- Synthesis Of Purine Nucleotides • Atoms of purine ring are contributed by: Asp, Gly and Gln/ CO2/ and N10-formyl-tetrahydrofolate • Ring is constructed in the liver by reactions that add donated carbons and nitrogens to preformed ribose 5-phosphate. Figure 22.5Sources of individual atoms in the purine ring
A. Synthesis of 5-phosphoribosyl-1-pyrophosphate (PRPP) • PRPP is an “activated pentose”, participates in synthesis of purines and pyrimidines, and in salvage of purine bases • Synthesis of PRPP from ATP and ribose-5-P is catalyzed by PRPP synthetase (ribose phosphate pyrophosphokinase). • The enzyme is activated by the inorganic phosphate (Pi) and inhibited by purine nucleotides (end-product) • Sugar in PRPP is ribose, ribonucleotides are the end products of de novo purine synthesis • When deoxyribonucleotides needed for DNA syhtnesis, ribose is reduced
Figure 22.6Synthesis of 5-phosphoribosyl-1-pyrophosphate (PRPP), showing the activator and inhibitors of the reaction.
B. Synthesis of 5`-phosphoribosylamine • Amide group of Glutamine replaces the pyrophosphate group attached to C1 of PRPP • The enzyme, glutamine: phosphoribosyl pyrophosphate amidotransferase, is inhibited by purine 5` nucleotides AMP, GMP (end products). • This is the committed step in purine nucleotide biosynthesis • The rate of reaction is also controlled by Glutamine and PRPP conc. (intracellular PRPP conc. is normally far below the Km for the amidotransferase, i.e., small changes in [PRPP] cause proportional change)
C. Synthesis of inosine monophosphate, the “parent” purine nucleotide • The next 9 steps in purine nucleotide biosynthesis lead to the synthesis of Inosine Monophosphate (IMP, whose base is hypoxanthine). • The pathway requires 4 ATPs. • Two steps require N10-formyltetrahydrofolate.
Figure 22.7 Synthesis of purine nucleotides, showing the inhibitory effect of some structural analogs.
D. Synthetic inhibitors of purine synthesis • Sulfonamides, synthetic inhibitors, inhibit growth of rapidly dividing microorganisms without interfering with human cell functions. • Other purine synthesis inhibitors (eg. Methotrexate, structural analog of folic acid) used to control spread of cancer by interfering with synthesis of nucleotides (thus DNA, RNA). • Trimethoprim (another folate analog) has antibacterial activity as it selectively inhibits bacterial dihydrofolate reductase.
D. Synthetic inhibitors of purine synthesis • Inhibitors of human purine synthesis are extremely toxic to tissues, especially to developing structures such as in a fetus , or to cell types that replicate rapidly, including those of BM, skin, GI tract, immune system, or hair follicles. • Individuals taking such anti-cancer drugs experience adverse effects e.g., anemia, scaly skin, GI tract disturbance, immunodeficiencies, and baldness
E. Conversion of IMP to AMP and GMP • This requires a 2-step energy requiring pathway • The synthesis of AMP requires guanosine triphosphate (GTP) as an energy source, whereas the synthesis of GMP requires ATP • The 1st reaction in each pathway is inhibited by the end product. • This diverts IMP to the synthesis of the purine species present in lesser amounts. • If both AMP and GMP present in adequate amounts, de novo pathway of purine synthesis is turned off at amidotransferase step.
Figure 22.8 Conversion of IMP to AMP and GMP showing feedback inhibition.
F. Conversion of nucleoside monophosphates to nucleoside diphosphates and triphosphates • Nucleoside diphosphates (NDPs) are synthesized from Nucleoside monophosphates (NMPs) by base-specificnucleoside monophosphate kinases • These kinases do not discriminate between ribose or deoxyribose in the substrate. • ATP is generally the source of transferred phosphate as it is the abundant Nucleoside Triphosphate (NTP) • Adenylate kinase is particularly active in liver and muscle, where turnover of energy from ATP is high. • Its function is to maintain an equilibrium among AMP, ADP and ATP • NDPs and NTPs are interconverted by nucleoside diphosphate kinase, an enzyme with broad specificity.
Figure 22.9 Conversion of nucleoside monophosphates to nucleoside diphosphates and triphosphates.
G. Salvage pathway of purines • Purines from normal turnover of cellular nucleic acids, or obtained from diet and not degraded, can be reconverted into NTPs and used by the body • This is referred to as the “salvage pathway” of purines. • Conversion of purine bases to nucleotides: • Two enzymes are involved: • Adenine phosphoribosyltransferase (APRT) and • Hypoxanthine-guanine phosphoribosyltransferase (HPRT) • Both enzymes use PRPP as source of ribose 5-p. • The release of pyrophospahte (ppi) makes these reactions irreversible. • Adenosine is the only purine nucleoside to be salvaged
2. Lesch-Nyhan syndrome: • X-linked, recessive disorder, associated with virtually complete deficiency of HPRT. • Inability to salvage hypoxanthine or guanine, from which excessive amounts of uric acid are produced. • In addition, lack of salvage pathway causes increased PRPP levels and decreased IMP and GMP levels. • So, glutamine:phosphoribosylpyrophosphate amidotransferase has excess substrate and decreased inhibitors available, and de novo purine synthesis increased. • Decreased purine reutilization and increased purine synthesis results in production of large amounts of uric acid, making Lesch-Nyhan syndrome a severe, heritable form of gout. • Patients with Lesch-Nyhan syndrome tend to produce urate kidney stones. • In addition, characteristic neurologic features of the disorder include self-mutilation (biting of lips and fingers) and involuntary movements.
4. Synthesis of Deoxyribonucleotides • The nucleotides describes thus far contain ribose • The nucleotides required for DNA synthesis, however, are 2'-deoxyribonucleotides, which are produced from ribonucleoside diphosphates by the enzyme ribonucleotide reductase • The same enzyme acts on pyrimidine ribonucleotides.
A- Ribonucleotide reductase • It is a multi-subunit enzyme composed of two nonidentical dimeric subunits, 2B1 and 2B2 • Specific for the reduction of nucleoside diphosphates (ADP, GDP, CDP, UDP) to their deoxy forms. • The immediate donors of hydrogen atoms needed for reduction of 2`-OH are two –SH groups on the enzyme itself, which during reaction form a disulfide bond.
Figure 22.12 Conversion of ribonucleotides to deoxyribonucleotides.
A- Ribonucleotide reductase 1. Regeneration of reduced enzyme. • In order for ribonucleotide reductase to continue to produce deoxyribonucleotides, the disulfide bond created during the production of the 2'-deoxy carbon must be reduced • The source of the reducing equivalents for this purpose is thioredoxin—a peptide coenzyme of ribonucleotide reductase • Thioredoxin contains two cysteine residues separated by two amino acids in the peptide chain. • The two –SH groups of thioredoxin donate their H atoms to the enzyme, in the process forming S-S bond. 2. Regeneration of reduced thioredoxin. • The necessary reducing equivalents are provided by NADPH & H+, and the reaction is catalyzed by thioredoxinreductase (see Figure 22.12)
B. Regulation of deoxyribonucleotide synthesis • Ribonucleotide reductase is responsible for maintaining a balanced supply of the deoxyribonucleotides required for DNA synthesis. • Regulation of the enzyme is complex. • In addition to the single catalytic (active) site, there are allosteric sites on the enzyme involved in regulating its activity. • Activity sites: • The binding of dATP to allosteric sites (known as the activity sites) on the enzyme inhibits the overall catalytic activity of the enzyme and therefore prevents reduction of any of the four NDPs. • This effectively prevents DNA synthesis, and explains the toxicity of increased levels of dATP seen in conditions such as adenosine deaminase deficiency. • In contrast, ATP bound to these sites activates the enzyme.
B. Regulation of deoxyribonucleotide synthesis • Substrate specificity sites: • The binding of nucleoside triphosphates to additional allosteric sites (known as the substrate specificity sites) on the enzyme regulates substrate specificity, causing an increase in the conversion of different species of ribonucleotides to deoxyribonucleotides as they are required for DNA synthesis. • E.g., deoxythymidinetriphosphate (dTTP) binding at the specificity sites causes a conformational change that allows reduction of GDP to dGDP at the catalytic site.
The drug, hydroxyurea destroys the free radical required for enzymatic activity of ribonucleotide reductase, and thus inhibits the generation of substrates for DNA synthesis. • Hydroxyurea has been used in the treatment of cancers such as chronic myelogenous leukemia. • Hydroxyurea is also used in the treatment of sickle cell disease however, the increase in fetal hemoglobin seen with hydroxyurea has not been linked to its effect on ribonucleotide reductase.