Download

1 / 86
930 likes | 1.34k Vues
Biomolecules: Lipids and Their Metabolism. Lipids are naturally occurring organic molecules isolated from organisms by extraction with nonpolar organic solvents Fats, oils, waxes, some vitamins and hormones, and most nonprotein cell-membrane components Lipids have limited solubility in water

E N D
Biomolecules: Lipids and Their Metabolism Lipids are naturally occurring organic molecules isolated from organisms by extraction with nonpolar organic solvents • Fats, oils, waxes, some vitamins and hormones, and most nonprotein cell-membrane components • Lipids have limited solubility in water • Lipids defined by their physical property (solubility) and not by their structure
Biomolecules: Lipids and Their Metabolism Lipids classified into two broad types: • Those that contain ester linkages that can be hydrolyzed • Include fats and waxes • Steroids which do not have ester linkages and cannot be hydrolyzed • Includes cholesterol and other steroidal hormones
23.1 Waxes, Fats, and Oils Waxes are mixtures of esters of long-chain carboxylic acids with long-chain alcohols • Carboxylic acid usually has an even number of carbons from 24 through 36 • Beeswax is the ester of the C30 alcohol triacontan-1-ol and the C16 acid hexadecanoic acid
Waxes, Fats, and Oils Animal fats and vegetable oils are the most widely occurring lipids • Closely related structures • Have different appearances • Animal fats are solids like butter and lard • Vegetable oils are liquids like corn and peanut oil
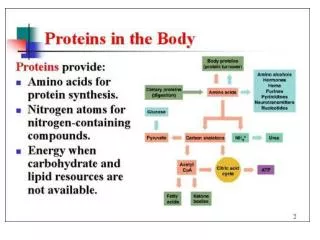
Waxes, Fats, and Oils • Fats and oils are triglycerides, or triacylglycerols • Triesters of glycerol with three long-chain carboxylic acids called fatty acids • Fats are used by animals for long-term energy storage • Less highly oxidized than carbohydrates • Provide about six times as much energy as an equal weight of glycogen
Waxes, Fats, and Oils Hydrolysis of a fat or oil with aqueous NaOH yields glycerol and three fatty acids • Fatty acids are generally unbranched and contain an even number of carbon atoms between 12 and 20 • If double bonds are present they are generally Z, or cis • The three fatty acids of a specific triacylglycerol do not need to be the same • A fat or oil is likely to be a complex mixture of many different triacyglycerols
Waxes, Fats, and Oils Structures of some common fatty acids
Waxes, Fats, and Oils Approximate composition of some fats and oils
Waxes, Fats, and Oils Palmitic acid (C16) and stearic acid (C18) are the most abundant saturated fatty acids Oleic and linoleic acids (both C18) are the most abundant unsaturated fatty acids
Waxes, Fats, and Oils • Oleic acid is monounsaturated • It has only one double bond • Linoleic , linolenic, and arachidonic acids are polyunsaturated fatty acids – they contain more than one double bond • Linoleic and linolenic acids occur in cream and are essential to the human diet • Infants grow poorly and develop skin lesions if fed a diet of nonfat milk for prolonged periods
Waxes, Fats, and Oils Unsaturated fatty acids generally have lower melting points than their saturated counterparts • Vegetable oils have higher proportions of unsaturated to saturated fatty acids and have lower melting points • Unsaturated fatty acids have C=C double bonds that make crystal formation more difficult thus lowering the melting point • Saturated fatty acids have a uniform shape that allows them to pack together efficiently in a crystal lattice thus raising their melting point
Waxes, Fats, and Oils Unsaturated fatty acids can be catalytically hydrogenated • Catalytic hydrogenation is typically carried out at high temperature using a nickel catalyst • Margarine and shortening are produced by hydrogenating soybean, peanut, or cottonseed oil until the proper consistency is obtained • Hydrogenation is accompanied with cis-trans isomerization of the double bonds • Produces fats with about 10% to 15% trans unsaturated fatty acids • Dietary intake of trans fatty acids increases cholesterol levels in the blood increasing the risk of coronary artery disease
Waxes, Fats, and Oils The conversion of linoleic acid into elaidic acid
23.2 Soap Soap has been known since at least 600 BC • Phoenicians prepared a curdy material by boiling goat fat with extracts of wood ash • Wood ash was used as a source of alkali until the early 1800s when Na2CO3 was made by heating sodium sulfate with limestone • Cleansing properties of soap were not generally recognized until the 18th century • Soap is a mixture of sodium or potassium salts of long-chain fatty acids produced by hydrolysis (saponification) of animal fat with alkali
Soap Crude soaps are purified by adding NaCl or KCl to precipitate the pure carboxylate salts • Smooth soap that precipitates is dried, perfumed, and pressed into bars for household use • Soaps act as cleansers because the two ends of a soap molecule are different • The carboxylate end of the long-chain molecule is ionic and therefore hydrophilic • The long hydrocarbon portion of the molecule is nonpolar and hydrophobic • The net effect of these two opposing tendencies is that soaps are attracted to both oils and water and therefore useful as cleansers
Soap When soaps are dispersed in water they form micelles, molecular spheres with both hydrophobic and hydrophilic properties • The long hydrophobic tails cluster together on the hydrophobic inside of the spheres • The ionic heads group together on the hydrophilic surface of the spheres out into the water • Grease and oil droplets are solubilized in water when they are coated by the nonpolar tails of soap molecules in the center of the micelles
Soap Soluble sodium carboxylates are converted by Mg2+, Ca2+, and Fe3+ found in hard water to insoluble metal salts • Mg2+, Ca2+, and Fe3+ carboxylate salts of fatty acids constitute soap scum • Synthetic detergents based on salts of long-chain alkylbenzenesulfonic acids (sulfonate detergents) do not form insoluble metal salts in hard water • The alkylbenzene end is attracted to grease • The anionic end is attracted to water Sulfonate detergents do not form insoluble metal salts in hard water and do not leave an unpleasant scum.
23.3 Phospholipids Phospholipids are diesters of phosphoric acid, H3PO4
Phospholipids Two general kinds of phospholipids • Glycerophospholipids • Based on phosphatidic acid • Acyl group at C1 is usually saturated and the one at C2 is usually unsaturated • The phosphate group at C3 is also bonded to an amino alcohol such as choline, ethanolamine, or serine • Compounds are chiral at C2 with an L or R configuration
Phospholipids • Sphingomyelins • Have sphingosine or a related dihydroxyamine as their backbone • Abundant in brain and nerve tissue • Major constituent of the coating around nerve fibers
Phospholipids Phospholipids occur widely in both plant and animal tissues • Constitute approximately 50% to 60% of cell membranes • Organize in cell membrane into a lipid bilayer about 5.0 nm thick • Nonpolar tails aggregate in the center of the bilayer • Bilayer serves as barrier to passage of water, ions, and other components into and out or cells
23.4 Catabolism of Triacylglycerols: The Fate of Glycerol Triacyglycerol catabolism begins with hydrolysis to yield glycerol plus fatty acids • Reaction catalyzed by lipase • Aspartic acid, histidine, and serine residues in enzyme’s active site catalyze individual steps • Hydrolysis involves two sequential nucleophilic acyl substitution reactions • One covalently binds an acyl group to the side-chain –OH group of a serine residue on the enzyme • One frees the fatty acid from the enzyme
Catabolism of Triacylglycerols: The Fate of Glycerol Figure 23.3 Mechanism of action of lipase
Catabolism of Triacylglycerols: The Fate of Glycerol STEPS 1-2 OF FIGURE 23.3: ACYL ENZYME FORMATION • Deprotonation of serine alcohol by histidine • Deprotonated serine adds to a carbonyl group of a triacylglycerol to give a tetrahedral intermediate • The tetrahedral intermediate expels a diacylglycerol as the leaving group and produces an acyl enzyme
Catabolism of Triacylglycerols: The Fate of Glycerol STEPS 3-4 OF FIGURE 23.3: HYDROLYSIS • Water is deprotonated by histidine • HO- adds to enzyme-bound acyl group • Tetrahedral intermediate expels neutral serine residue as the leaving group freeing the fatty acid • Enzyme is returned to its active form
Catabolism of Triacylglycerols: The Fate of Glycerol Fatty acids released on triacylglycerol hydrolysis are transported to mitochondria and degraded to acetyl CoA • Glycerol is transported to liver for further metabolism • In the liver glycerol is phosphorylated by reaction with ATP and then oxidized by NAD+ • C2 of glycerol is prochiral • Enzyme selects pro-R arm for reaction • Resultant dihydroxyacetone phosphate (DHAP) enters the carbohydrate glycolysis pathway
23.5 Catabolism of Triacylglycerols: β- Oxidation Figure 23.4 Fatty acyl CoA’s are catabolized by repetitive four-step sequence known as b-oxidation pathway • Pathway results in cleavage of an acetyl group from end of fatty-acid chain • Chain-shortening step is a retro-Claisen reaction of a b-keto thioester
Catabolism of Triacylglycerols: β-Oxidation STEP 1 OF FIGURE 23.4: INTRODUCTION OF A DOUBLE BOND b-Oxidation pathway begins with removal of hydrogen atoms from C2 and C3 of fatty acyl CoA • Introduction of double bond into carbonyl usually involves coenzyme flavin adenine dinucleotide (FAD) • Reduced FADH2 is the by-product
Catabolism of Triacylglycerols: β-Oxidation Exact mechanism of FAD-catalyzed reaction is unknown • Can be either two-electron (polar) or one-electron (radical) pathway • First step involves abstraction of pro-R hydrogen from a position of acyl CoA giving thioester enolate ion • Pro-R hydrogen at b position is transferred to FAD • Resultanta,b-unsaturated acyl CoA has trans double bond
Catabolism of Triacylglycerols: β-Oxidation Mechanism of step one of b-oxidation may occur by hydride-transfer • Electrons on enolate ion might expel a b hydride ion which could add to doubly bonded N5 nitrogen on FAD • Protonation of intermediate at N1 gives product
Catabolism of Triacylglycerols: β-Oxidation STEP 2 OF FIGURE 23.4: CONJUGATE ADDITION OF WATER a,b-Unsaturated acyl CoA reacts with water by a conjugate addition pathway to yield a b-hydroxyacyl CoA • Process catalyzed by enoyl-CoA hydratase • Intermediate thioester enolate ion is protonated by addition of hydrogen to the a position
Catabolism of Triacylglycerols: β-Oxidation STEP 3 OF FIGURE 23.4: ALCOHOL OXIDATION b-Hydroxyacyl CoA is oxidized to a b-ketoacyl CoA • Reaction catalyzed by L-3-hydroxyacyl-CoA dehydrogenase • Oxidation requires NAD+ as a coenzyme • Reaction yields NADH/H+ as by-product
Catabolism of Triacylglycerols: β-Oxidation STEP 4 OF FIGURE 23.4: CHAIN CLEAVAGE Acetyl CoA is split off from chain leaving an acyl CoA that is two carbon atoms shorter than original • Reaction is catalyzed by b-ketoacyl-CoA thiolase • Mechanistically the reverse of a Claisen condensation reaction
Catabolism of Triacylglycerols: β-Oxidation • Reaction occurs by nucleophilic addition of a cysteine –SH group in the active site of the enzyme to the keto group of the b-ketoacyl CoA • Cleavage of C2-C3 bond followed by expulsion of acetyl CoA enolate ion • Enzyme-bound acyl group undergoes nucleophilic acyl substitution by reaction with coenzyme A • Resultant chain-shortened acyl CoA enters another b-oxidation round for further degradation
Catabolism of Triacylglycerols: β-Oxidation Figure 23.5 Catabolism of the 14-carbon myristic acid by the b-oxidation pathway yields seven molecules of acetyl CoA after six passages
Catabolism of Triacylglycerols: β-Oxidation Most fatty acids have even numbers of carbon atoms • None is left over after b-oxidation • Fatty acids with an odd number of carbon atoms yield the three-carbon propionyl CoA in the final oxidation • Propionyl CoA is converted to succinate which enters the citric acid cycle
23.6 Biosynthesis of Fatty Acids All fatty acids are derived biosynthetically from acetyl CoA by sequential addition of two-carbon units • Acetyl CoA arises primarily from metabolic breakdown of carbohydrates in glycolysis • Dietary carbohydrates consumed in excess of immediate energy needs are turned into fats for storage • Fatty-acid synthesis in vertebrates is catalyzed by a large, multienzyme complex containing two identical subunits of 2505 amino acids each
Biosynthesis of Fatty Acids Figure 23.5 Pathway for fatty-acid biosynthesis from the two-carbon precursor acetyl CoA
Biosynthesis of Fatty Acids STEPS 1-2 OF FIGURE 23.6: ACYL TRANSFERS Thioester acetyl CoA, the ultimate product of glycolysis, is the starting material for fatty-acid biosynthesis • Primingreactions render acetyl CoA more reactive • First priming reaction is a nucleophilic acyl substitution that converts acetyl CoA into acetyl ACP (acyl carrier protein) • Reaction catalyzed by ACP transacylase
Biosynthesis of Fatty Acids • Acyl group in acetyl ACP is linked by a thioester bond to the sulfur atom of phosphopantetheine • Phosphopantetheine is linked to ACP through the side-chain –OH group of a serine residue • Second priming reaction involves further exchange of thioester linkages by another nucleophilic acyl substitution • Results in covalent bonding of acetyl group to a cysteine residue in the synthase complex • Catalyzes subsequent condensation step
Biosynthesis of Fatty Acids STEPS 3-4 OF FIGURE 23.6: CARBOXYLATION AND ACYL TRANSFER • Third step is a loading reaction • Acetyl CoA is carboxylated by reaction with HCO3- and ATP to yield malonyl CoA plus ADP • Coenzyme biotin needed as carrier of CO2 • Fourth step binds malonyl group to ACP arm of multienzyme synthase
Biosynthesis of Fatty Acids STEP 5 OF FIGURE 23.6: CONDENSATION Claisen condensation is key carbon-carbon bond-forming reaction that builds fatty-acid chain • Acetyl synthase is the electrophilic acceptor and malonyl ACP is the nucleophilic donor • Mechanism may involve decarboxylation of malonyl ACP to give enolate followed by immediate addition of the enolate ion to the carbonyl group of acetyl synthase • Breakdown of tetrahedral intermediate gives 4-carbon condensation product acetoacetyl ACP
Biosynthesis of Fatty Acids STEPS 6-8 OF FIGURE 23.6: REDUCTION AND DEHYDRATION Ketone carbonyl group in acetoacetyl ACP is next reduced to the alcohol b-hydroxybutyryl ACP by b-keto thioester reductase and NADPH • Results in R stereochemistry at newly formed chirality center in b-hydroxy thioester product
Biosynthesis of Fatty Acids Subsequent dehydration of b-hydroxybutyryl ACP by an E1cB reaction in step 7 yields trans-crotonyl ACP • Carbon-carbon double bond of crotonyl ACP is reduced by NADPH in step 8 to yield butyryl ACP • Double-bond reduction occurs by conjugate addition of hydride ion from NADPH to b-carbon of trans-crotonyl ACP
Biosynthesis of Fatty Acids Net effect of eight steps in fatty-acid biosynthesis pathway is to take two 2-carbon acetyl groups and combine them into a 4-carbon butyryl group Further condensation of the butyryl group with another malonyl ACP yields a 6-carbon unit, and so forth Chain elongation of 16-carbon palmitic acid occurs by similar but not identical reactions with CoA acting as the carrier group
23.7 Terpenoids Terpenoids are a vast and diverse group of lipids found in all living organisms • All contain a multiple of five carbons • All derived biosynthetically from the five-carbon precursor isopentenyl diphosphate • Terpenoids contain oxygen • Terpenes are hydrocarbons
Terpenoids Monoterpenoids • Contain 10 carbons • Derived from two isopentenyl diphosphates • Found primarily in plants, bacteria, and fungi Sesquiterpenoids • Contain 15 carbons • Derived from three isopentenyl diphosphates • Found primarily in plants, bacteria, and fungi Diterpenoids • Contain 20 carbons • Derived from four isopentenyl diphosphates • Found in both plants and animals Higher terpenoids • Triterpenoid lanosterol (C30) • Precursor for steroid hormones • Tetraterpenoid b-carotene (C40) is dietary source of vitamin A
Terpenoids Terpenoid precursor isopentenyl diphosphate • Formerly called isopentenyl pyrophosphate • Abbreviated IPP • Biosynthesized by two different pathways • In animals and higher plants sesquiterpenoids and triterpenoids arise primarily from the mevalonate pathway
Terpenoids Figure 23.9 The Mevalonate Pathway to Isopentenyl Diphosphate • Begins with conversion of acetate to acetyl CoA • Claisen condensation yields acetoacetyl CoA • Second Claisen condensation reaction with third molecule of acetyl CoA yields six-carbon compound 3-hydroxy-3-methylglutaryl CoA • 3-Hydroxy-3-methylglutaryl CoA is reduced to mevalonate • Phosphorylation with loss of CO2 and phosphate ion completes synthesis