Download

1 / 32
350 likes | 623 Vues
GABA A and GABA B Receptor Abnormalities in Autism. S. Hossein Fatemi, M.D., Ph.D. Professor of Psychiatry, Pharmacology and Neuroscience University of Minnesota Medical School Minneapolis, MN, USA. Autism. Neurodevelopmental disorder

E N D
S. Hossein Fatemi, M.D., Ph.D. Professor of Psychiatry, Pharmacology and Neuroscience University of Minnesota Medical School Minneapolis, MN, USA
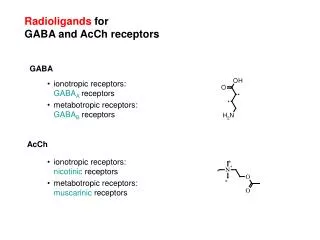
Autism • Neurodevelopmental disorder • Brain pathology involving parietal, frontal, cerebellar, hippocampal, and amygdalar areas • Early onset of disease • Genetic etiologies • MZ concordance 36-95% • Chromosomes 1, 5, 6, 8, 13, 15, 16, 18, 21, x, y • HOX A, Reelin, Wnt • Environmental etiologies • Rubella, borna, syphilis, CMV, mumps • Thalidomide • Valproic acid • Mercury
Brain Pathology in Autism • Selective cortical atrophy • Purkinje cell loss and atrophy (Piven; Fatemi) • Smaller cingulate gyrus • Higher packing density and smaller neuron size in hippocampus, amygdala and septal nucleus (Bauman and Kemper) • Macrocephaly (Courchesne) • Abnormal brain stem • Microcolumnar pathology (Casanova et al.)
Pathological Findings • Cell migration – Reelin • GABA transmission – GAD • Apoptosis – Bcl2 and P53 • Cell injury – GFAP • cAMP/cGMP modulation - PDEs
Methods • SDS-PAGE • Western Blotting • qRT-PCR • HPLC
Table 1. Demographic data for subjects with autism and controls Dx, diagnosis; PMI, postmortem interval; M, male; F, female; Autistic*, all autistic subjects were also mentally retarded[2]; MVA, motor vehicle accident
Figure 1. Representative samples of GABRA1 (51 kDa), GABRA2 (51 kDa), GABRA3 (55 kDa), GABRA5 (52 kDa), GABRA6 (50 kDa), GABRG2 (45 kDa), GABRG3 (51 kDa), GABRB3 (56 kDa), GABBR1 (108 kDa), GABBR2 (105 kDa), and β-Actin (42 kDa) in BA9, BA40, and cerebellum of subjects with autism (A) and matched controls (C).
Table 2. Expression of GABRA1-GABRA3, GABRB3, GABBR1, and GABBR2 in cerebellum, BA40, and BA9 in subjects with autism vs. controls *, Two-tailed independent group t-tests; **, p<0.05; nc, no change
Table 3. Expression of GABRA5, GABRA6, GABRG2, and GABRG3 in cerebellum, BA40, and BA9 in subjects with autism vs. controls *, Two-tailed independent group t-tests; **, p<0.05; nc, no change
Figure 2. Neurochemical analysis of brain regions of control and autistic subjects in BA9 (A), BA40 (B), and cerebellum (C). Shown are mean values ± SEM of the mean for dopamine (DA), serotonin (5-HT), glutamate (Glu), glutamine (Gln), taurine (tau), and GABA. Statistical significance of difference analyzed by one way analysis of variance followed by Holm Sidak post hoc test. **p<0.05
Protein Levels • GABAAa1 protein by 63% in cerebellum • GABAAb3 protein by 51% in cerebellum • GABAB R1 protein by 67% in cerebellum • GABAB R2 protein by 46% in cerebellum • GABAAa1 protein by 52% in BA40 • GABAAa2 protein by 39% in BA40 • GABAAa3 protein by 57% in BA40 • GABAAb3 protein by 38% in BA40 • GABAB R1 protein by 71% in BA40 • GABAAa1 protein by 65% in BA9 • GABAB R1 protein by 70% in BA9
HPLC Studies • cerebellar serotonin (p<0.05) in autism • No difference in levels of GABA, glutamate, dopamine, glycine, or taurine between two group • Supported by central serotonergic hypofunction in autism (Croonenbergh et al., 2007)
mRNA Levels • mRNA for GABAAb3 levels by 29% in cerebella of autistics vs. controls • mRNA for GABAB R1 levels by 15% in cerebella of autistics vs. controls
GABAAb3 • Implicated by multiple association studies in autism (Buxbaum JD et al., 2002; Cook EH et al., 1998; McCauley JL et al., 2004; Shao Y et al., 2003) • expression in autism, Angelman syndrome and Rett’s disease (Samaco RC et al., 2005) • Pentameric chloride channels • Crucial site of action for intravenous anesthetics, ethanol, and developmental processes (i.e., ventromedial nucleus of hypothalamus) • Localized to 15q11-q13, a site for b3, a5, and g3 subunits of GABAA receptor • Our results are the first to show significant decreases in b3 proteins in cerebellum and BA40 of autistics, as well as b3 mRNA and protein decreases in cerebellum of autistics
GABAB R1 • Signal transduction/G protein activation (Jones KA et al., 2000) • Downregulation in kainic acid-induced seizures in rats signify neurodegeneration (Furtingers et al., 2003) • Impairment of attentional processing (Prosser HM et al., 2001) • Ca-dependent receptor activation; tubulin-dependent receptor trafficking • Insulin resistance, hyperlocomotion, and atypical absence seizure • Our results are the first to demonstrate R1 deficiency in 3 important brain areas in autism
GABAB R2 • Gene locus at 9q22.1 • G-protein coupled receptor 51 • Associated with nicotine dependence • Increased with absence seizure in rat somatosensory cortex and thalamus (Principalle et al., 2003)
GABAAa1 • Gene locus 5q34-q35 • Associated with juvenile myoclonic epilepsy • Associated with depression in probands (Murray et al., 1994) • 5HT1A receptor in brains of subjects with JME (Meschaks et al., 2005). Serotonin is reduced in cerebellum of autistics (Fatemi et al., 2008) • No previous publication indicating its reduction in autism
GABAAa2 • Gene locus at 4p12 • Modulates anxiety and stress response • May be involved in plasticity of ventrobasal complex and posterior nucleus of thalamus in chronic inflammatory pain (Ferreria-Gomes et al., 2006) • Expression in medial amygdala (Byrnes et al., 2007) • Involved in hippocampal dentate granule cell during development (Brooks-Kayal et al., 2001) • Associated with alcohol dependence (Soyka et al., 2008)
GABAAa3 • Gene locus at Xq28 • Localized to substantia nigra parvalbumin-positive nonpigmented cells (Waldvogel et al., 2008) • Involved in unipolar major depression (Henket et al., 2004)
GABAAa5 • Gene locus at 15q11.2-q12 • Component of a pentameric receptor mediating inhibitory neurotransmission • Involved in the susceptibility to schizophrenia (Papadimitriou et al., 2001a) and bipolar affective disorder (Papadimitriou et al., 2001b; Otani et al., 2005) • Implicated in autism (Menold et al., 2001; Ashley-Koch et al., 2005; Tochigi et al., 2007), Angelman syndrome (Nurmi et al., 2001), and childhood absence epilepsy (CAE) (Lu et al., 2004)
GABAAa6 • Gene locus at 5q34 • Component of a pentameric receptor mediating inhibitory neurotransmission, expressed in the cerebellar granule cells and the related cells of cochlear nucleus • Implicated in schizophrenia (Petryshen et al., 2005), alcoholism (Chang et al., 2002; Dick et al., 2003; Sen et al., 2004), and heroin abuse (Loh et al., 2007)
GABAAg2 • Gene locus at 5q31.1-q33.1 • This subunit carries the benzodiazepine binding site • Component of a pentameric receptor mediating inhibitory neurotransmission, complexing with DRD5 and promoting mutually inhibitory functional interactions between these receptor systems • Implicated in schizophrenia (Petryshen et al., 2005) and mood disorders (Yamada, 2003) • Involved in the physiological dependence on alcohol (Sander et al., 1999; Chang et al., 2002; Dick et al., 2003), heroin abuse (Loh et al., 2007), and methamphetamine abuse (Nishiyama et al., 2005) • Cause of childhood absence epilepsy type 2 (ECA2) (Olsen et al., 1999; Wallace et al., 2001) generalized epilepsy with febrile seizures plus type 3 (GEFS+3) (Baulac et al., 2001) severe myoclonic epilepsy in infancy (SMEI)(Jansen et al., 2006)
GABAAg3 • Gene locus at 15q12 • This subunit also carries the benzodiazepine binding site • Component of a pentameric receptor mediating inhibitory neurotransmission • Associated with alcohol dependence (Dick et al., 2004) • Associated with risk for autism (Ma et al., 2005; Ashley-Koch et al., 2006)
Glutamic Acid Decarboxylase 65kDa • Localized to axon terminals • Membrane bound and in vesicles • Involved in vesicular release of GABA • Involved in synthesis of GABA by phasically firing neurons • Can exist as either amphiphillic GAD65 homodimer or GAD65/67 heterodimer
Glutamic Acid Decarboxylase 67kDa • Concentrated in interneurons • Cytosolic • Involved in non-vesicular synthesis of GABA • Involved in continuous synthesis of GABA in tonically firing neurons • Used for synthesis of GABA for general metabolic activity • Can exist as either hydrophillic GAD67 homodimer or amphiphillic GAD65/67 homodimer
Glutamate & aspartate in plasma of autistic children (Moreno et al., 1992; Moreno-Fuenmayer et al., 1996) • CSF glutamate in 4 patients with Rett’s syndrome (Hamberger et al., 1992) • Glutamate & GABA in platelets of drug-naïve autistic patients (Rolf et al., 1993) • Glutamate to N-acetylaspartate ratio in gray matter in Rett’s syndrome (Pan et al., 1999) • mRNA of excitatory amino acid transporter 1 and AMPA1 receptor with density of AMPA1 receptor in autistic cerebellum (Purcell et al., 2001)
Conclusions • GABAAa1 and b3, GABAB R1 and R2 proteins decreased in cerebellum of autistics (p<0.05) • GABAAa1, a2, a3, and b3 and GABAB R1 proteins decreased in BA40 of autistics (p<0.05) • GABAAa1, a5, a6, g2, and GABAB R1 proteins decreased in BA9 of autistics (p<0.05) • GABAAb3 and GABAB R1 mRNAs decreased in cerebellum of autistics (p<0.05) • Serotonin levels reduced in cerebellum of autistics without any change in levels of glutamate or GABA • Results support ↑ rates of seizure disorder and mental retardation in autistic subjects
Acknowledgements Human tissue was obtained from the NICHD Brain and Tissue Bank for Developmental Disorders; the Harvard Brain Tissue Resource Center, which is supported in part by PHS grant number R24 MH068855; the Brain Endowment Bank, which is funded in part by the National Parkinson Foundation, Inc., Miami, Florida; and the Autism Tissue Program and is gratefully acknowledged. Grant support by National Institute of Child Health and Human Development (#5R01HD052074-01A2) to SHF is also gratefully acknowledged.
Collaborators • University of Minnesota, Minneapolis, MN USA - TJ Reutiman, TD Folsom, PD Thuras • Charité Campus Mitte, University Medicine, Berlin, Germany - C Winter, R Sohr, J Klein • University of Rochester, Rochester, NY USA - DA Pearce, M Zanche