Hemoglobin (haemoglobin, Hb, Hgb)
170 likes | 382 Vues
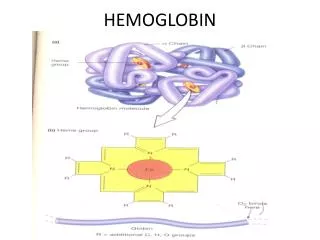
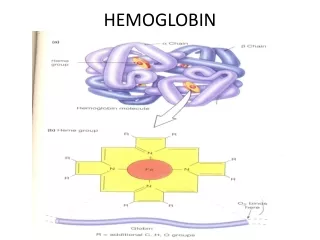
Hemoglobin (haemoglobin, Hb, Hgb). red cells in blood carry O 2 from lung to tissues by hemoglobin, a 4-subunit protein having an O 2 -binding prosthetic group, heme, that gives blood its color (Hb also carries (some) CO 2 and H + back to the lung)

Hemoglobin (haemoglobin, Hb, Hgb)
E N D
Presentation Transcript
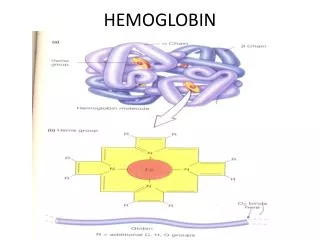
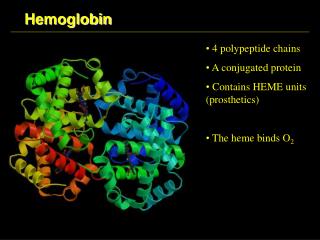
Hemoglobin (haemoglobin, Hb, Hgb) red cells in blood carry O2 from lung to tissues by hemoglobin, a 4-subunit protein having an O2-binding prosthetic group, heme, that gives blood its color (Hb also carries (some) CO2 and H+ back to the lung) one of the first proteins the structure of which has been determined for (Perutz, 1959;Noble Prize in Chemistry shared with Kendrew in 1962) contrary to Hb, myoglobin (Mb) is a monomeric O2-storage molecule, evolutionary related to Hb, avery similar structure to Hb (globin fold) Hb can use 90% of its full O2-binding potential, while Mb can use only 7% in Hb there is a so-called cooperativity, so O2-binding to one chain potentiates O2-binding to the rest of the 3 chains(same true for release) H+- and CO2-binding to Hb also modulate O2-binding the hypothesis that alteration in amino acid sequence may lead to disease was first proven with Hb and Mb (sickle-cell anemia; single mutation in Hb)
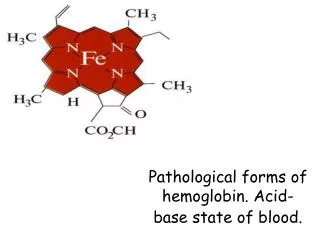
O2-free or O2-bound states exist: deoxy-Hb/Mb or oxy-Hb/Mb high a-helical content connected by b-turns in Hb/Mb with one O2-binding site/monomer on each heme prosthetic group under normal conditions, heme binds Fe2+, there are a 5th and a 6th coordi- nation site for Fe on each side of the heme plane the 5th coordination of Fe takes place with a His side-schain (imidazole) of the protein (proximal His); the 6th site is unoccupied in the deoxy-pro- tein (ready for binding O2) Fe is too big for the hole in the middle of the porphyrin ring (deoxy-Mb/Hb)
upon oxygenation: rearranged electon-structure of Fe upon O2-binding results in a slightly smaller size that now fits in the middle of the tetrapyrrole ring(magnetic properties also change for Hb and that is the basis of functional magnetic resonance imaging, fMRI, one of the most powerful methods of examining brain function) Linus Pauling predicted this as well in 1936, 25 years ahead of time
O2-bound Fe acts rather as a Fe3+- O2- (superoxide anion) complex (charge- transfer complex, mixture of resonance structures);it is crucial to release oxygen as O2 because superoxideis a reactive oxygen species (ROS) and can generate further harmful species that damage various cellular compo- nents (proteins, DNA, lipids, membranes, etc.) and this would leave Fe as Fe3+ that with heme would constitute for metmyoglobin/methemoglobin that does not bind O2 (O2-storage capacity is lost)…structural features of Mb/Hb stabilize the O2-Hb/Mb complex in such a way that will assure that oxygen will be released as O2 there is another His (distal His) that donates a H-bond to the bound O2 (the superoxide character of bound O2 strengthens this interaction; the protein part of Hb/Mb controls O2-binding and release) Mb’s P1/2 value (50% of saturation of available binding sites) for O2 is at 2 torr (Hg mm), simple saturation curve
Hb`sO2-binding curve lookslike an “S” letter called a sigmoid curve (signi- ficantly weaker O2-binding than for Mb at the same O2-tension, P1/2=26 torr; this is true in red blood cells where Hb binds a special molecule, 2,3-bis- phosphoglycerate, as well, that lowers Hb`s affinity to O2 significantly)
Physiological importance of sigmoidal O2-binding in the lungs partial pressure of O2 is high (100 torr, 98% of Hb binds O2), in actively metabolizing tissues it is 20 torr (32% saturation; in resting state 40 torr); 98-32=66% of the binding sites contribute to O2-transport (these numbers would be 98%vs. 91% for Mb, only 7% difference – too small window to work with, Mb binds O2 too tight for O2-transport) these numbers would be for a protein with theoretically optimal affinity for O2, 63% and 25% (38% difference), cooperativity is the best solution for delivering O2 to tissues (~10x better than it would be possible with Mb)
100 torr (lung) to 40 torr (resting muscle, 60 torr drop in partial pressure) means 98% to 77%=21% drop in O2-saturation of Hb 40 torr (resting muscle) to 20 torr (in exercise, 20 torr drop) means 77% to 32%=45% drop in O2-binding of Hb steepest part of the O2-binding curve is right there where the most O2 is needed, so when we switch from resting to exercise
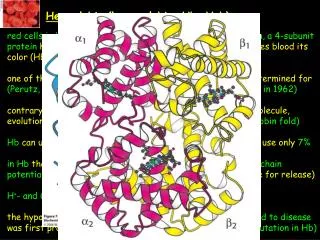
How is cooperativity delivered at the atomic level? O2-binding sites are far away from each other, direct interaction is not possible upon O2-binding the a1-b1 and a2-b2 dimers rotate ~15o with respect to one another (substantial conformational change, the connecting interface changes the most, the dimers themselves not that much, the dimers are freer to move and O2-binding sites are free of strain and are capable of binding O2 with a much higher affinity in the oxygenated state) Tense state, deoxy-Hb, constrained by subunit- subunit interactions Relaxed state, fully oxygenated form
Concerted or MWC model: only two states exist, T and R states, in the R state O2-binding is much stronger, occupying more and more binding sites with O2 shifts the equi- librium towards the R state Sequential model: binding of a ligand changes the conformation of the subunit that it binds to, which consequently will change the conformation of a neighboring subunit so that its affinity for O2 would be increased and so on neither model is fully describing the actual mechanism of action of Hb
How does O2-binding at one site shift the T to R equilibrium for the tetramer? upon O2-binding Fe moves inplane which pulls also on proximal His proximal His is part of an a-helix that also moves with the His the C-terminus of this helix lies in the interface of the two ab dimers and the movement of this C-terminal region triggers the T-to-R transformation of Hb the structural transition at the iron ion in one of the subunits is directly transmitted to the other subunits (the rearrangement of the inter-dimer interface makes communication between subunits possible enabling the cooperative binding of O2
2,3-BPG and the O2-affinity of Hb pure Hb binds O2 much tighter than Hb in red cells and the reason is the presence of 2,3-BPG in red cells (~2mM, just as much as Hb itself) without 2,3-BPG binding to Hb, Hb would be able to release only8% of its O2-load in tissues crystal structure of deoxy-Hb bound to 2,3-BPG reveals that a single mole- cule of 2,3-BPG binds to a Hb tetramer in a pocket in the center of the tetramer; this pocket exists only in the T-form and gets collapsed in the R- form (and 2,3-BPG gets released) in order to make the T-to-R transition happen 2,3-BPG must dissociate off of Hb in the presence of 2,3-BPG more O2-binding sites must be occupied to induce the T-to-R transition, hence Hb stays in the lower affinity T state until higher O2 concentrations are available 2,3-BPG is an allosteric effector (see later)
Fetal Hb fetal Hb consists of 2 a and 2 g chains (latter has 72% sequence homo- logy to the b Hb chain) H143S is an important change (mutation) as H143 is part of the 2,3-BPG binding site; this mutation removes positive charges from the binding pocket lowering the affinity of 2,3-BPG to this site consequently fetal Hb will bind O2 with higher affinity than maternal Hb elegant solution from nature to solve an important biological problem (to transport O2 from mother to fetus)
The Bohr effect rapidly metabolizing tissues such as contracting muscle generate a lot of H+ and CO2 Hb is able to release more O2 where need augments (e.g. in contracting muscle) by the action ofH+ and CO2(also allosteric effectors; Bohr effect) lowering pH lowers O2-affinity of Hb (from 7.4 (lung) to 7.2 (tissue) with an 80 torr drop in pressure this amounts to a 77% (rather than 66% at pH=7.4) total carrying capacity) the mechanism of action is revealed, but not discussed here (rather complex mechanism) CO2 action is also partly due to acidification (carbonic acid for- mation)
reaction with N-terminus (switch of charge) carbamate termini participate in salt-bridge interactions in the ab inter-dimer interface and stabilize the T-state (lower O2-affinity, also 14% of CO2-transport)
Sickle-Cell Anemia (SCA) large fibrous aggregates of Hb, distorted red cells that clog small capilla- ries and impair normal blood flow (symptoms: painful swelling of extremities, higher risk for stroke, anemia; 1% of West Africans suffer from this dise- ase) E6V mutation is the responsible in the b chain, the mutant Hb is called HbS, deoxy-HbS has a very low solubility, both alleles are affected in disease Why Africa? people heterozygous for SCA are resistant to malaria