Download

1 / 19
190 likes | 211 Vues
Explore how plants extract energy from the environment through processes like respiration and photosynthesis. Learn about the importance of bioenergetics in governing terrestrial life and the significance of thermodynamic laws in biological systems. Discover how ATP serves as the energy currency of living cells and the role of redox reactions in maintaining cellular energy balance.

E N D
Plant cell bioenergetics Plant Physiology and Biotechnology Lecture 2 Chapter 2 Jan. 5, 2006
Plant and animal life strategies differ • Life = resistance to thermodynamic laws • Need energy to fight __________ • How to extract energy from the environment? Adopt one of two metabolic strategies: be either of… (Chemi)heterotroph • Ingest complex organic molecules • Use RESPIRATION to make energy while oxidizing these molecules to simple molecules (Photo)autotroph • Use the sun’s energy to make complex molecules out of simple molecules • Use respiration to oxidize the complex molecules to _____ and ______ . OR
Bioenergetics govern terrestrial life • All forms of life on earth are dependent on the ability of plants to perform photosynthesis • Humans are at the top of the food chain and consume plants directly or indirectly (herbivores) explorelearning.com • Bioenergetics is the study of how organisms use and make energy through • Photosynthesis and • Respiration These processes capture energy to do work. • The flow of energy is governed by thermodynamic laws
Bioenergetics are in turn dependent on the Thermodynamic Laws We learn these in physics class, but they have special relevance in biology Law 1: The energy of the universe is constant • Energy not lost in reactions, merely passes to different forms Law 2: The capacity of an isolated system to perform useful work constantly decreases • Systems increase in entropy – this is energy that is not available to do work • Glucose (C6H12O6) has less entropy than CO2
Environmental energy may be harnesssed to do work • Energy that is available to do work is Gibbs free energy, or G H = G + TS H=total energy G=energy available for work TS=energy unavailable for work (entropy) • Individual chemical reactions have more or less ability to do work • Quantify them from their change in G or ΔG • Negative ΔG = spontaneous, will occur without input of energy • This means that • G products < G reactants • Reactions with a negative ΔG can also be said to be • Exergonic = energy yielding • Exothermic = generate heat
Reactions with a negative Gibbs free energy occur spontaneously • The chemical reactions involved in catalysis of complex molecules (e.g. glucose) usually have a negativeΔG • Reactions that have a positive ΔG will not occur spontaneously e.g., 6CO2 + 6H2O → C6H12O6 + 6H2O • If G changes, so does S • not all energy is useable for work → ENTROPY always increases • Biochemists have a special definition of ΔG • The standard free energy change, or ΔG°’, defined at • pH 7.0 • Conc. of reactants and products both at 1 M
The mass action ratio is a measure of whether a chemical reaction occurs spontaneously • Let’s consider a chemical reaction S → P • Left alone to come to equilibrium, the mass action ratio Keq is a measure of the concentrations of substrates and products under any given condition: Keq = [P]eq / [S]eq ←MASS ACTION RATIO • If the concentrations of S and P are displaced away from equilibrium, free energy (G) is available to do work as the reaction proceeds towards equilibrium → free energy changes → ΔGº′
How do you make chemical reactions with +ΔG occur in vivo? • Couple them to exergonic reactions • Net free energy change (ΔG) must be negative for coupled chemical reactions to proceed to product formation ATP → ADP + PiΔGº′ = -32.2 kJ/mol Glucose + Pi → G6P+13.8 Net: Glc + Pi + ATP → G6P +ADP + Pi-18.4
Maintenance of high ATP levels is a hallmark of living cells • ATP hydrolysis is the energy currency of the cell (high energy molecule) • The Keq of the ATP ADP + Pi reaction in vitro is 105 • Recall that Keq is a measure of [P]/[S] • This means ATP hydroysis is very spontaneous! • Contrast this to the situation in vivo: photosynthesis and respiration maintain a high cellular concentration of ATP • The Keq of the ATP ADP + Pi reaction = 10-4 in vivo • 109 difference in Keqbetween the reaction in vitro and in vivo! • As ATP is hydrolysed to ADP, this exothermic reaction can do a lot of work! • LIVING CELLS: • Maintain a non-equilibrium steady-state • A high [ATP] allows plant cells to be energetically homeostatic
Photosynthesis and respiration are a series of chemical reactions • Intermediates in these pathways are sequentially oxidized (loses H) and reduced (gains H) • The operation of these pathways is dependent on maintaining reducing power to drive the individual reactions forward • Reducing power possesses negative ΔGº′ that can be used to drive endergonic reactions towards product formation • Reducing power involves passing electrons and protons between molecules • It is thus sensitive to pH in vivo! • Photosynthesis and respiration allow the cell to control e- flow in a predictable way and profit energetically from this flow • e.g. in electron transport chains of • Mitochondria: oxidative phosphorylation • Chloroplasts: phostosynthetic electron transport • PUTTING IT ALL TOGETHER: How does transport of electrons and protons control ATP synthesis? • Mitochondria and chloroplasts link e- transport to ATP synthesis
Chemiosmosis links electron transport and ATP synthesis The chemiosmotic theory was proposed by Peter Mitchell in 1961 • Energy transducing membranes are impermeable to H+ (inner membranes in chloroplasts and mitochondria) • e- carriers are arranged asymmetrically in membrane • Some e- carriers act as proton pumps • Import H+ against the H+ gradient • Preserve some energy of e- transport
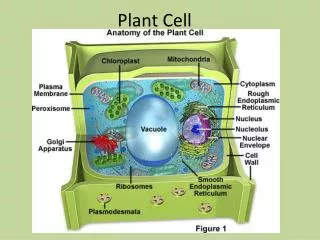
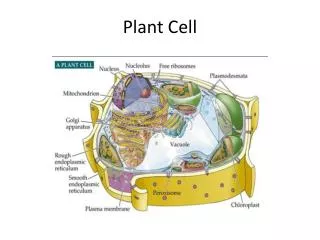
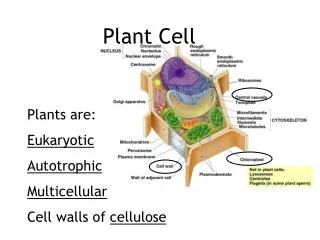
Chloroplast and mitochondrial structure permit their role in ATP synthesis • Energy transduction is possible because mitochondria and chloroplasts have different compartments • These compartments make it possible to sequester ion gradients that drive ATP synthesis Fig. 2.6 • Chloroplasts are only found in photosynthetic tissue
Chloroplasts possess a complex internal structure • STROMA: • unstructured background matrix • Lots of protein: PCR/Calvin cycle enzymes (dark reactions) • DNA • THYLAKOIDS: • Membranes stacked as grana (tightly oppressed thylakoids) enclosing LUMEN • can connect between grana Fig. 2.5 • Membranes contain • Pigments (chlorophyll, carotenoids) • Photosynthetic E.T.C. • LUMEN: • Site of H2O oxidation: H2O → 2H+ + O2 • Reservoir for H+
Mitochondria and chloroplasts both perform chemiosmosis • MITOCHONDRIA are similarly structured to chloroplasts • Double membrane, outer freely permeable, inner selectively permeable • Inner membrane: impermeable to H+, metabolites • LOTS OF PROTEIN • Invaginations: • CRISTAE: increased surface area for ATP synthesis • MATRIX: filled with enzymes involved in C metabolism, mtDNA • There is a higher number of mitochondria in metabolically active cells • Synthesize ATP via RESPIRATION (oxidative phosphorylation) Fig. 1.20
Chloroplasts and mitochondria both pump protons to make ATP Fig. 2.6c • Similar structure, similar function • Both synthesize ATP by chemiosmosis • Electron transport in membrane imports H+ inside lumen / intermembrane space • This causes proton motive force via • pH gradient and thus an • Electrochemical gradient • This gradient creates a large amount of potential chemical energy • Powered by large -ΔG of photosynthetic E.T.C. • Hard to hold H+ inside lumen want to diffuse back into stroma • BUT: thylakoid membrane is impermeable to H+
Chemiosmosis links pH gradients to ATP synthesis in both mitochondria and chloroplasts • H+ must pass through large protein complex (multimeric) to pass lumen → stroma • ATP synthase! • In chloroplast: light dependent ATP synthesis (uses H2O → O2) called PHOTOPHOSPHORYLATION • In mitochondria: chemiosmotic ATP synthesis (O2: terminal e- acceptor, makes H2O) called OXIDATIVE PHOSPHORYLATION We will discuss the specifics of chloroplast versus mitochondrial electron transport in lectures on photosynthesis Fig. 2.7: chloroplast ATP synthase
Experimental proof that the proton gradient creates ATP Light increases stromal pH because protons are being transferred into the lumen by the chloroplast electron transport chain • Therefore, plant chloroplasts use light (the photosynthetic E.T.C.) to establish the H+ gradient • Can also do this on isolated thylakoids • Make ATP in the DARK! • This is proof that the pH gradient drives the synthesis of ATP Lights off, the H+ leak slowly back into the stroma through the thylakoid membrane Simulate this effect in the dark by changing the pH ! “Stroma” Lumen Use the chemical potential of the gradient to make ATP Establish pH gradient Fig. 2.8: making ATP in the dark
ATP synthesis is reversible • We have seen how establishment of a H+ gradient is necessary for ATP synthesis in both mitochondria and chloroplasts ATP synthesis is also reversible! • ATP synthase and similar proteins that span the thylakoid membrane can also hydrolyse ATP back to ADP + Pi • Use energy (-ΔGº′) to drive H+ back into lumen • ATP synthases are also ATPases: also important for ion transport across thylakoid membrane
Proton gradients make ATP • What does the cell do with this ATP? • Powers growth and development • Can also make ATP molecule by molecule in chemical reactions of metabolism using enzymes: proteins that are biological catalysts • a/k/a substrate-level phosphorylation • e.g. pyruvate kinase glycolysis ADP ATP TCA cycle PEP pyruvate malate Pyruvate kinase Knowledge of bioenergetics is a necessary prerequisite for understantding future lectures on photosynthesis, water transport, carbon metabolism, germination, growth and development