Download

1 / 85
900 likes | 1.22k Vues
第五章 基因表达的调控 ( Regulation and control of gene expression ). 概 述. 一、基因表达的概念. 基因表达 ( gene expression). 生物基因组中结构基因所携带的遗传信息,经过转录、翻译等一系列过程,合成具有特定的生物学功能和生物学效应的蛋白质的全过程。. (一)时间特异性 (temporal specificity) 按功能需要某一特定基因表达严格按特定的时间顺序发生。. 二、基因表达的时间性及空间性. (二)空间特异性 (spatial specificity).

E N D
第五章基因表达的调控(Regulation and control of gene expression)
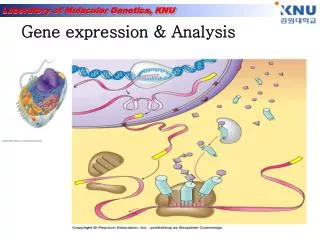
一、基因表达的概念 基因表达( gene expression) 生物基因组中结构基因所携带的遗传信息,经过转录、翻译等一系列过程,合成具有特定的生物学功能和生物学效应的蛋白质的全过程。
(一)时间特异性 (temporal specificity) 按功能需要某一特定基因表达严格按特定的时间顺序发生。 二、基因表达的时间性及空间性
(二)空间特异性(spatial specificity) 指在个体生长全过程,某种基因产物在个体不同组织器官表达存在差异。又称细胞或组织特异性(cell or tissue specificity)。
三、基因表达的方式 按对刺激的反应性分为两大类: (一)基本(组成性)表达 (constitutive gene expression) 基因较少受环境因素影响,而是在个体各个生长阶段的大多数或几乎全部组织中持续表达,或变化很小。如管家基因。
(二)诱导和阻遏表达 诱导表达(induction expression) 在特定环境信号刺激下,有些基因的表达表现为开放或增强。 阻遏表达(repression expression) 在特定环境信号刺激下,有些基因的表达表现为关闭或下降。
协调表达(coordinance expression) 在一定机制下,功能相关的一组基因,协调一致,共同表达。
机体各种细胞中含有的相同遗传信息(相同的结构基因),根据机体的不同发育阶段、不同的组织细胞及不同的功能状态,选择性、程序性地表达特定数量的特定基因的过程。 四、基因表达的调控的概念
蛋白质 ( 主要 ) 五、基因表达的调控因子: • 小分子RNA ( 某些环节 )
基因组 六、基因表达的调控水平 • 转录 • 转录后 • 翻译 • 翻译后
原核生物基因表达调控主要在转录水平,其次是翻译水平。原核生物基因表达调控主要在转录水平,其次是翻译水平。 本节主要以大肠杆菌为例介绍原核生物基因表达的调控。 第一节原核生物基因表达的调控
1.只有一种RNA聚合酶。 2.基因表达以操纵子为基本单位。 一、原核生物基因表达的特点 3.转录和翻译偶联进行。 4.mRNA翻译起始部位有特殊的碱基序列----SD序列。
在起始密码AUG上游4~13个碱基之前有一段富含嘌呤的序列,其一致序列(consensus sequence)为AGGAGG,能与核糖体30S亚基中16S rRNA 3′端富含嘧啶的序列结合,与蛋白质合成过程中起始复合物生成有关。 SD( Shine-Dalgarno )序列
5.原核生物基因表达的调控主要在转录水平,即对RNA合成的调控。5.原核生物基因表达的调控主要在转录水平,即对RNA合成的调控。 通常有两种方式: (1) 起始调控,即启动子调控; (2) 终止调控,即衰减子调控。
(一)转录起始的调控 1.б因子及RNA 聚合酶与转录起始的调控 (1) б因子调控RNA 聚合酶与DNA结合: 确保RNA 聚合酶与特异启动区稳定结合,而不是与其它位点结合。 二、原核生物基因表达的调控机制
(2)RNA 聚合酶与启动区序列的相互作用 RNA 聚合酶:大肠杆菌仅有的一种RNA 聚合酶,要识别相当数量的启动区,需依赖数目繁多的辅助蛋白(如б因子)完成这些功能。 启动区序列:保守序列
启动序列 是RNA聚合酶结合并启动转录的特异DNA序列。 -35区 -10区 RNA转录起始 trp TTGACA N17 TTAACT N7 A A tRNATyr TTTACA N16 TATGAT N7 lac TTTACA N17 TATGTT N6 A recA N16 TATAAT N7 A TTGATA Ara BAD CTGACG N16 TACTGT N6 A 共有序列 TTGACA TATAAT
共有序列决定启动序列的转录活性大小。 某些特异因子(蛋白质)决定RNA聚合酶对一个或一套启动序列的特异性识别和结合能力。
操纵子(operon):一个多顺反子转录单位与其调控序列即构成操纵子。操纵子(operon):一个多顺反子转录单位与其调控序列即构成操纵子。 2.转录起始的负调控 乳糖操纵子(lactose operon,lac)是原核生物基因转录负调控的最典型模式。
调控区 结构基因 P O Z Y A Z: β-半乳糖苷酶 Y: 透酶 A:乙酰基转移酶 操纵元件 阻遏基因I 启动子 CAP结合位点 乳糖操纵子(lac operon)的结构 DNA
仅有葡萄糖 葡萄糖耗完,有乳糖存在
-10 +1 +10 +20 +30 . . . . . 5′ATGTTGTGTGGAATTGTGAGCGGATAACAATTTCACACAGGAA 3′ 3′TACAACACACCTTAACACTCGCCTATTGTTAAAGTGTGTCCTT 5′ lac操纵区序列是一个以+11位GC碱基对为中心的反向重复序列 与lac阻遏蛋白结合的操纵基因区域具有反向重复序列,这种反向重复序列是能与蛋白质特异结合的DNA的特征性结构。
阿拉伯糖操纵子是正调控的典型例子。 3.转录起始的正调控 阿拉伯糖操纵子的基因结构图 操纵子由结构基因B、A、D以及调控元件I1、I2、O1、O2和启动子构成。araC基因编码调节蛋白AraC。
AraC 对阿拉伯糖操纵子的调节图 不存在阿拉伯糖时,AraC二聚体与O1、O2及I1结合,二聚体间相互作用使DNA弯曲成环结构。由于I2不被占据,B、A、D基因不发生转录,但有低水平的araC基因转录。
AraC 对阿拉伯糖操纵子的调节图 当有阿拉伯糖时,AraC二聚体与阿拉伯糖形成复合物,使Ara C形状改变,I1和I2由一个AraC二聚体占据,从而激发B、A、D基因转录。此时,可有一过性高水平的araC mRNA形成,维持到O1-O2环结构形成。
在大肠杆菌的许多操纵子中,基因的转录不是由单一因子调控的,而是通过负调控因子和正调控因子进行复合调控。典型例子:糖代谢有关的操纵子,如lac操纵子。在大肠杆菌的许多操纵子中,基因的转录不是由单一因子调控的,而是通过负调控因子和正调控因子进行复合调控。典型例子:糖代谢有关的操纵子,如lac操纵子。 4.转录起始的复合调控
cAMP:环一磷酸腺苷 CAP:分解代谢基因激活蛋白质 (catabolite gene activator protein,CAP) cAMP与葡萄糖相关性:葡萄糖↑,cAMP↓ 葡萄糖↓,cAMP↑ CAP的活性依赖cAMP, 形成cAMP-CAP复合 物,促进多种操纵子的转录起始。
CAP CAP CAP CAP CAP CAP + + + +转录 DNA P O Z Y A 无葡萄糖,cAMP浓度高时 有葡萄糖,cAMP浓度低时
RNA-pol O O mRNA O O 有乳糖 没有乳糖 葡萄糖低 cAMP浓度高 葡萄糖高cAMP浓度低
※当阻遏蛋白封闭转录时,CAP对该系统不能发挥作用;※当阻遏蛋白封闭转录时,CAP对该系统不能发挥作用; ※如无CAP存在,即使没有阻遏蛋白与操纵序列结合,操纵子仍无转录活性。
原核生物的转录终止调控方式分两大类:A.依赖ρ因子的终止调控;B.不依赖ρ因子的终止调控。此外,核糖体也参与转录终止。原核生物的转录终止调控方式分两大类:A.依赖ρ因子的终止调控;B.不依赖ρ因子的终止调控。此外,核糖体也参与转录终止。 (二)转录终止的调控 例:色氨酸操纵子表达的调控有两种方式: a.通过阻遏蛋白的负调控 b.通过衰减子作用
Trp (2)转录衰减的调控 调节区 结构基因 P O RNA聚合酶 trpR RNA聚合酶 Trp 低时 mRNA 6700个核苷酸 Trp 高时 ? 色氨酸操纵子 140个核苷酸
结构基因 调节区 P O trpR 前导mRNA 1 2 3 4 UUUU…… 终止密码子 trp 密码子 衰减子区域 衰减子结构 UUUU…… UUUU…… UUUU…… 前导序列 14aa前导肽编码区: 包含序列1 第10、11密码子为trp密码子 形成发夹结构能力: 序列1/2>序列2/3>序列3/4
色氨酸操纵子mRNA引导序列不同区域互补所形成的不同二级结构色氨酸操纵子mRNA引导序列不同区域互补所形成的不同二级结构 引导序列由一段14氨基酸前导肽编码区和一个衰减子组成,AUG密码子后面紧跟13个密码子,第10、11为色氨酸密码子。
色氨酸操纵子的转录衰减作用 色氨酸丰富时,核蛋白体顺利沿引导序列移动直达最后一个密码子UGA,合成完整的引导肽。RNA 聚合酶停止在衰减子部位。 色氨酸缺乏时,核蛋白体终止在1区Trp密码子部位,RNA 聚合酶通过衰减子而继续转录。
意义:这种机制可以保证充分地消耗色氨酸,使其合成维持在满足需要的水平,防止色氨酸堆积和过多地消耗能量。同时,也使细菌能够优先将环境中的色氨酸消耗完,然后开始自身合成。意义:这种机制可以保证充分地消耗色氨酸,使其合成维持在满足需要的水平,防止色氨酸堆积和过多地消耗能量。同时,也使细菌能够优先将环境中的色氨酸消耗完,然后开始自身合成。 39
(三) 翻译水平的调控 翻译一般在起始和终止阶段受到调节。 调节分子:RNA、蛋白质 调节分子可直接或间接决定翻译起始位点能否为核蛋白体所利用。
反义RNA 指能与特定mRNA互补结合的RNA片段,反义RNA由反义基因转录而来。天然的具有功能的反义RNA分子一般在200个碱基以下。反义RNA又称mRNA干扰性互补RNA(mRNA interfering complementary RNA, micRNA)。 1.反义RNA的调控作用 41
①反义RNA与mRNA 5ˊ端非翻译区包括SD序列相结合,阻止mRNA与核糖体小亚基结合,直接抑制翻译。 反义RNA有三种作用方式: ②反义RNA与mRNA 5ˊ端编码区起始密码子AUG结合,抑制mRNA翻译起始。 ③反义RNA与mRNA的非编码区互补结合,使mRNA构象改变,影响其与核糖体结合,间接抑制了mRNA的翻译。 42
反义RNA调控Tn10转位酶基因的表达示意图 Tn10转位酶基因由PIN启动子控制,反义RNA的基因由POUT启动子控制。两个启动子方向相反,转录区有35个碱基重叠,互补区覆盖了Tn10转位酶mRNA的翻译起始区,活性POUT比PIN更强。结果是转位酶的表达效率非常低,转位作用极少发生。细菌DNA突变机会减少,易于存活。 43
mRNA的降解速度是翻译调控的另一个重要机制。蛋白质合成速率的快速改变,与许多细菌mRNA降解速度很快有很大关系。 例:E.coli的许多mRNA在37℃时的平均寿命大约为2min,细菌可以对环境变化作出快速反应。 2. RNA的稳定性与调控的关系
3.蛋白质合成中的自身调控 大肠杆菌核糖体蛋白有50余种,它们既可与 rRNA 组装成核糖体,也可与自身 mRNA 翻译起始区结合。50种核糖体蛋白的基因分布在几个不同的操纵子中。
第二节 真核生物基因表达的调控 单细胞真核生物,如酵母基因表达的调控和原核生物表达的调控基本相同。 多细胞真核生物,遗传信息从细胞核的基因组DNA传递到基因编码的蛋白质均受到多层次的调控。
(1)细胞具有全能性全能性:指同一种生物的所有细胞都含有相同的基因组DNA,都有发育成完整个体的潜能。 一、真核生物基因表达的特点 (2)基因表达的时间性和空间性时间性:个体发育的不同阶段,基因表达的种类和数量是不同的;空间性:在不同组织和器官中,基因表达的种类和数量是不同的。
(3)转录和翻译分开进行 真核生物DNA在核中转录生成 mRNA,在胞质中合成蛋白质。 (4)初级转录产物要经过转录后加工修饰 初级转录产物——核不均一性RNA(heterogeneous nuclear RNA ,hnRNA),要经过戴帽(m7GpppN)、加PolyA尾、切除插入的内含子和拼接外显子等过程,才形成成熟的mRNA分子。
(5)不存在超基因式操纵子结构 真核生物基因转录产物为单顺反子,一条 mRNA只翻译一种蛋白质。功能相关的基因大多数分散在不同的染色体上,分别进行转录。 (6)部分基因多拷贝 既可满足细胞的需要,也是表达调控的一种有效方式。
二、真核生物基因表达调控的机制 (一)基因组DNA水平的调控 1.染色质的丢失 不可逆的调控。如一些低等生物(如线虫)体细胞发育过程中发生染色质丢失。
2.基因扩增(gene amplification) 当细胞对某种基因产物需要量剧增,单纯靠调节其表达活性不足满足需要,只有增加这种基因的拷贝数。
3.基因重排(gene rearrangement) 指某些基因片段改变原来存在顺序而重新排列组合,成为一个完整的转录单位。调节表达产物多样性。