Protein Structure and Prediction
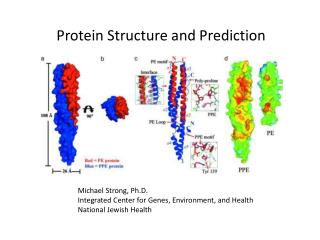
Protein Structure and Prediction. Michael Strong, Ph.D. Integrated Center for Genes, Environment, and Health National Jewish Health. From Sequence to Structure. HIV Protease With Inhibitor. HIV Protease. Experimental Approach.

Protein Structure and Prediction
E N D
Presentation Transcript
Protein Structure and Prediction Michael Strong, Ph.D. Integrated Center for Genes, Environment, and Health National Jewish Health
From Sequence to Structure HIV Protease With Inhibitor HIV Protease Experimental Approach PQITLWKRPLVTIRIGGQLKEALLDTGADDTVLEEMNLPGKWKPKMIGGIGGFIKVRQYDQIPIEICGHKAIGTVLVGPT PVNIIGRNLLTQIGCTLNF
From Sequence to Structure H1N1 NA MNPNQKIITIGSVCMTIGMANLILQIGNIISIWISHSIQLGNQN QIETCNQSVITYENNTWVNQTYVNISNTNFAAGQSVVSVKLAGNSSLCPVSGWAIYSK DNSVRIGSKGDVFVIREPFISCSPLECRTFFLTQGALLNDKHSNGTIKDRSPYRTLMS CPIGEVPSPYNSRFESVAWSASACHDGINWLTIGISGPDNGAVAVLKYNGIITDTIKS WRNNILRTQESECACVNGSCFTVMTDGPSNGQASYKIFRIEKGKIVKSVEMNAPNYHY EECSCYPDSSEITCVCRDNWHGSNRPWVSFNQNLEYQIGYICSGIFGDNPRPNDKTGS CGPVSSNGANGVKGFSFKYGNGVWIGRTKSISSRNGFEMIWDPNGWTGTDNNFSIKQD IVGINEWSGYSGSFVQHPELTGLDCIRPCFWVELIRGRPKENTIWTSGSSISFCGVNS DTVGWSWPDGAELPFTIDK" Computational Approach
Protein Building Blocks Typical Protein Sequence MNPNQKIITIGSVCMTIGMANLILQIGNIISIWISHSIQLGNQN
Amino Acid Side Chain (R groups) Disulfide Bonds
Amino Acid Side Chain (R groups) - acidic + basic
Most Proteins Spontaneously Fold DNA Transcribed by RNA polymerase RNA Translated by Ribosome Folded Protein Some proteins need chaperones for correct folding
native state, Folded protein spontaneous self-organisation (~1 second) Most Proteins Spontaneously Fold Folded protein Christian Anfinsen’s Experiment 1950s Denaturing conditions Unfolded protein Native conditions
Most Proteins Spontaneously Fold Important to Computational Biologists, because this suggests that all information relating to the correct folding of a protein is contained in it’s primary amino acid sequence, but …
Most Proteins Spontaneously Fold But Proteins lack easy rules for folding as compared to DNA DNA Protein
Many Factors Influence Protein Folding • Proteins Assume the Lowest Energy Structure • Factors that influence folding include: • Hydrophobic Interactions / collapse (particularly within the core) • Hydrogen bonds– lead to secondary structures • Disulfide Bonds (Cysteine residues) • Salt Bridges / Ionic Interactions (among charged residues) • Multimeric interactions with same type or other proteins Protein
Common Secondary Structures Alpha helix
Common Secondary Structures Beta Sheet
Common Secondary Structures Loop Regions Loop
Diversity of Protein Structures A B C D Streptomycin resistance gidB Homology model Pyrazinamide Activating enzyme pncA Crystal Structure Rifampin target rpoB Homology Model Fluoroquinolone Target gyrA Crystal Structure F G H E Streptomycin Resistance rpsL Homology model Ethionamide Target, inhA Crystal Structure Isoniazid Target inhA Crystal Structure Isoniazid Activating Enzyme, KatG Crystal Structure http://www.proteopedia.org/wiki/index.php/User:Michael_Strong/TB
Experimental Methods of Structure Determination X-ray crystallography High resolution structure determination Grow a protein Crystal
Experimental Methods of Structure Determination X-ray crystallography High resolution structure determination
Experimental Methods of Structure Determination X-ray crystallography High resolution structure determination • Intensities and phases of all reflections are combined in a Fouriertransform to provide • maps of electron density • Phases determined by using heavy metals or selenomethionine (MAD)
Experimental Methods of Structure Determination NMR – Nuclear Magnetic Resonance High resolution structure determination • Smaller Proteins than X-ray • Distances between pairs of hydrogen atoms • Lots of information about dynamics • Requires soluble, non-aggregating material • Assignment sometimes difficult NOE cross-peak if they are within5.0 Å
Experimental Methods of Structure Determination Cryo Electron Microscopy Low to medium resolution structure determination • Low to medium resolution ~10-15Å • Limited information about dynamics • Can be used for very large molecules and complexes
Database of Protein Structures PDB – Protein Data Bank
Database of Protein Structures PDB – Protein Data Bank 95,113 protein structures as of 10/31/2013
Database of Protein Structures PDB – Protein Data Bank Even so, the number of solved structures greatly lags behind the rate of new genes being sequenced … Solution: Computational Structural Methods
Database of Protein Structures PDB – Protein Data Bank Files • Atoms in pdb files are defined by their Cartesian coordinates:
Visualization of PDB files Pymol, Jmol, Chimera, etc
Visualization of PDB files Pymol, Jmol, Chimera, etc
DALI Structural Alignments Align Protein Structures, Structure Superposition Generates a comparison matrix (transform protein into a 2D array of distances between C-alpha atoms. Z score reflects reliability, lowest RMSD identified
From Sequence to Structure H1N1 NA MNPNQKIITIGSVCMTIGMANLILQIGNIISIWISHSIQLGNQN QIETCNQSVITYENNTWVNQTYVNISNTNFAAGQSVVSVKLAGNSSLCPVSGWAIYSK DNSVRIGSKGDVFVIREPFISCSPLECRTFFLTQGALLNDKHSNGTIKDRSPYRTLMS CPIGEVPSPYNSRFESVAWSASACHDGINWLTIGISGPDNGAVAVLKYNGIITDTIKS WRNNILRTQESECACVNGSCFTVMTDGPSNGQASYKIFRIEKGKIVKSVEMNAPNYHY EECSCYPDSSEITCVCRDNWHGSNRPWVSFNQNLEYQIGYICSGIFGDNPRPNDKTGS CGPVSSNGANGVKGFSFKYGNGVWIGRTKSISSRNGFEMIWDPNGWTGTDNNFSIKQD IVGINEWSGYSGSFVQHPELTGLDCIRPCFWVELIRGRPKENTIWTSGSSISFCGVNS DTVGWSWPDGAELPFTIDK" • Secondary Structure Prediction • Alpha Helix, Beta Strand, or Other • Tertiary Predictions: • Homology Modeling • Fold Recognition • De Novo Protein Structure Prediction Computational Approach
Secondary Structure Prediction 1st and 2nd generation – looked at probability of amino acid to be in a helix, strand, or other (coil/loop) based on known structures. Chou-Fasman (short runs of amino acids), GOR (Bayesian, takes neighbors into account) - helices – no prolines, periodicity 3.6 residues/turn - strands – alternating hydropathy, or ends hydrophillic and center hydrophobic -other – small, polar, flexible residues, and prolines But, stalled at 55- 60% accuracy 3rd generation – also used position specific profiles based on multiple sequence alignments (evolutionary information) (ie insertion/deletion more likely to be in coil/turn), PSI BLAST and HMM, NN and SVM (improved to about 75-80%)
Secondary Structure Prediction But we really want to know how the protein folds in three dimensions
But we really want to know how the protein folds in three dimensions
CASP - Critical Assessment of Techniques for Protein Structure Prediction • Started in 1994, Helped push the field of structure prediction • “Contest-like” setup • Catagories include: • Homology Modeling / Comparative Modeling • Fold Recognition / Threading • Ab Initio, De novo • Partially vs. Automated Methods (now quite similar results) Goal: Predict structures of solved but unpublished/unreleased structures (used to evaluate predictions. Every year, predictions / algorithms get better
Comparative Modeling “Homology Modeling” • Proteins that have similar sequences (i.e., related by evolution) are likely to have similar three-dimensional structures • 1. BLAST sequence of Interest against PDB to identify a template • Multiple templates can be used if desired • Templates with Ligands bound can be used to identify binding sites and interacting residues in the homology model Sequence identity required depends on protein length. A good rule of thumb is to have at least 40% sequence identity. Higher sequence identity is best. Lower than 25% is not reliable (zone of uncertainty) Above 75% sequence identity, usually quite reliable homology model Accurate sequence alignments very important Programs include Modeller and Swiss Model
Comparative Modeling “Homology Modeling” • Steps include: • Template recognition and initial alignment • Alignment Correction (Multiple Sequence Alignment can Help) • Backbone Generation (transfer coordinates from template) • Loop Modeling (loops hard to predict with insertions) • Side Chain Modeling (usually similar tortion angles at high sequenc ID) • Model Optimization (minor energy minimization steps or restrain some atom positions) • Model Validation (Higher ID more accurate usually, Calculate energy, or normality index (bond length, tortion angles)) • Iteration (to refine)
Protein Threading, Fold Recognition Often, seemingly unrelated proteins adopt similar folds. -Divergent evolution, convergent evolution. For sequences with low or no sequence homology Protein Threading § Generalization of homology modeling method • Homology Modeling: Align sequence to sequence • Threading: Align sequence to structure (templates) For each alignment, the probability that that each amino acid residue would occur in such an environment is calculated based on observed preferences in determined structures. § Rationale: • Limited number of basic folds found in nature • Amino acid preferences for different structural environments provides sufficient information to choose the best-fitting protein fold (structure)
Fold recognition 3.6 Å 5% ID NK-lysin (1nkl) Bacteriocin T102/as48 (1e68) • The number of possible protein structures/folds is limited (largenumber of sequences • but relatively few folds (some estimate ~1000)) (most apparent when 50% of structures with no seq homology were solved and had folds similar to known structures) 90% of new structures deposited in PDB have similar folds to those already known • Proteins that do not have similar sequences sometimes have similarthree-dimensionalstructures (such as B-barrel TIM fold) • A sequence whose structure is not known is fitted directly (or“threaded”) onto a known • structure and the “goodness of fit” isevaluated using a discriminatory function • Need ways to move model closer to the native structure
Ab initio prediction of protein structure – concept Difficult because search space is huge. Much larger conformational space Goal: Predict Structure only given its amino acid sequence In theory: Lowest Energy Conformation • Go from sequence to structure by sampling the conformational space ina reasonable • manner and select a native-like conformation using a gooddiscrimination function Difficult for sequences larger that 150aa Rosetta (David Baker lab) one of best (CASP evaluation)
Rosetta structure prediction • 2 phases • Low-resolution phase – statistical scoring function and fragment assembly • A. local structure conformations using info from PDB (3 and 9mer stretches) • B. multiple fragment substitution simulated annealing – to find best arrangement of the fragments (Monte Carlo Search) • C. low resolution ensemble of decoy conformations • 2. Atomic refinement phase using rotamers and small backbone angle moves(in populated regions of Ramachandran plot) • A. Refinement • B. Then structures clustered based on RMSD • C. Center of the Largest Clusters chosen as representative folds (likely to be correct fold)
Quality Assessment Ramachandran Plot – Phi Psi angles To identify residues that may be in wrong conformation Procheck, What_check