Enzyme Biosynthesis
Enzyme Biosynthesis. Tri Rini Nuringtyas. As we remember ! Most enzymes are proteins so Mechanism of enzyme synthesis is no different from protein synthesis in general The information which determines the primary sequence of an enzyme is contained in the order of DNA sequence.


Enzyme Biosynthesis
E N D
Presentation Transcript
Enzyme Biosynthesis Tri Rini Nuringtyas
As we remember ! Most enzymes are proteins so Mechanism of enzyme synthesis is no different from protein synthesis in general • The information which determines the primary sequence of an enzyme is contained in the order of DNA sequence
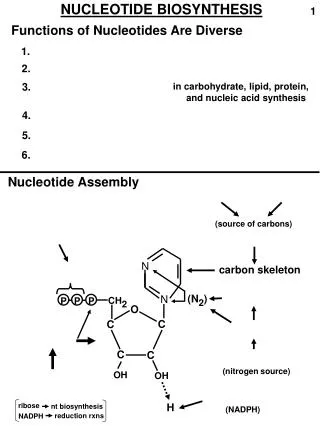
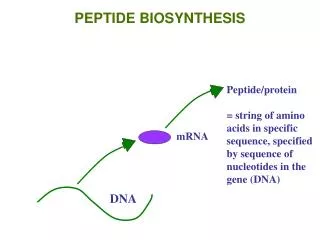
From gene to protein DNA Transcription mRNA Translation Sequence of a.a Primary structure of protein
Translation is the process of "reading" the codons and linking appropriate amino acids together through peptide bonds • Component of translation process • mRNA consist of genetic code • Ribosome • tRNA together with a.a • Enzymes
Translation process consists of 3 main stages • Initiation • Elongation • Termination Initiation Activation of amino acids for incorporation into proteins.
Genetic code Three nucleotides - codon - code for one amino acid in a protein Codon sequence of three nucleotides in a mRNA that specifies the incorporation of a specific amino acid into a protein. The relationship between codons and the amino acids they code for is called the genetic code.
Not all codons are used with equal frequency. There is a considerable amount of variation in the patterns of codon usage between different organisms.
Translation is accomplished by the anticodon loop of tRNA forming base pairs with the codon of mRNA in ribosomes
composed of a nucleic acid and a specific amino acid provide the link between the nucleic acid sequence of mRNA and the amino acid sequence it codes for. An anticodon a sequence of 3 nucleotides in a tRNA that is complementary to a codon of mRNA Transfer RNA (tRNA) Structure of tRNAs
Only tRNAfMetis accepted to form the initiation complex. • All further charged tRNAs require fully assembled (i.e., 70S) ribosomes • The Shine-Dalgarnosequence help ribosomes and mRNA aligns correctly for the start of translation. • Ribosome consists of • A site aminoacyl • P site peptidyl • - E site exit • Two initiation factors (IF1 &IF3) bind to a 70S ribosome. • promote the dissociation of 70S ribosomes into free 30S and 50S subunits. • mRNA and IF2, which carries • GTP • the charged tRNA • bind to a free 30S subunit. • After these have all bound, the 30S initiation complex is complete.
Peptide bond formation catalyzed by an enzyme complex called peptidyltransferase Peptidyltransferase consists of some ribosomal proteins and the ribosomal RNA acts as a ribozyme. The process is repeated until a termination signal is reached.
Termination of translationoccurs when one of the stop codons (UAA, UAG, or UGA) appears in the A site of the ribosome. No tRNAs correspond to those sequences, so no tRNA is bound during termination. Proteins called release factors participate in termination
Posttranslational Processing of Proteins • Folding • Amino acid modification (some proteins) • Proteolytic cleavage FOLDING Before a newly translated polypeptide can be active, it must be folded into the proper 3-D structure and it may have to associate with other subunits.
Enzymes/protein involve in folding process 1. Cis-trans isomerase for proline Proline is the only amino acid in proteins forms peptide bonds in which the trans isomer is only slightly favored (4 to 1 versus 1000 to 1 for other residues). Thus, during folding, there is a significant chance that the wrong proline isomer will form first. Cells have enzymes to catalyze the cis-trans isomerization necessary to speed correct folding. 2. disulfide bond making enzymes 3. Chaperonins (molecular chaperones) a protein to help keep it properly folded and non-aggregated.
Insulin is synthesized single polypeptide preproinsulin has leader sequence (help it be transported through the cell membrane) Specific protease cleaves leader sequence proinsulin. Proinsulin folds into specific 3D structure and disulfide bonds form Another protease cuts molecule insulin 2 polypeptide chains
Chaperones Function to keep a newly synthesized protein from either improperly folding or aggregating After synthesized, protein needs to fold in order to have its function The folding pattern is dictated in the amino acid sequence of the protein. • Some proteins capable to fold into its proper 3-D structure by itself without any help of other molecules • Some proteins need chaperones to fold (example in human hsp 70) • Some proteins need bigger protein chaperonins to be able to fold correctly. Chaperonins a polysubunit protein form “a cage” like shape give micro environment to protein
Protein Targeting Nascent proteins contain signal sequence determine their ultimate destination. Bacteria newly synthesized protein can: stay in the cytosol, send to the plasma membran, outer membrane, periplasmic, extracellular. Eukaryotes can direct proteins to internal sites lysosomes, mitochondria etc. Nascent polypeptide E.R and glycosylated golgi complex and modified sorted for delivery to lysosomes, secretory vesicle and plasma membrane.
Translocation • The protein to be translocated (called a pro-protein) is complexed in the cytoplasm with a chaperone • The complex keeps the protein from folding prematurely, which would prevent it from passing through the secretory porean ATPase that helps drive the translocation • after the pro-protein is translocated, the leader peptide is cleaved by a membrane-bound protease and the protein can fold into its active 3-d form.
Signal recognition particle (SRP) detects signal sequence and brings ribosome to the ER membrane
Most mitochondrial proteins are synthesized in the cytosol and imported into the organelle
Control of enzyme biosynthesis In living cell not all enzymes are synthesized with maximum velocity all the time. The rate of enzymes production controlled in accordance w/ • metabolic need • state of development of the cell The main point in the control of enzyme synthesis copying of the genes of the DNA in the form of mRNA
The inhibition of enzyme synthesis known as repression The operon model, as proposed in 1961 by Jacob and Monod.
A B The rates of formation of enzyme which are controlled by repressor regulated by the metabolic state of the cell [A] too high induction by substrate [B] too high repression by product Melibiose and IPTG a good inducer but not the substrate of galactose Gal repressor the action is prevented by the presence of D-galactose Repressor does not by itself bind to the operator has specific binding site for the product So repressor-product bind to the operator Example amino acid synthesis
Enzyme turn over Proteins are targeted for destruction Proteins have different half-lives Most enzymes that are important in metabolic regulation have short lives Also important for removal of abnormal proteins / enzymes Proteolytic enzymes are found through out the cell
Several proteases present in the eukaryotic cytosol two Ca2+ activated proteases calpains an ATP-dependent protease proteasome Four structural features are currently thought to be determinants of turnover rate : • Ubiquitination • Oxidation of amino acid residues • PEST sequences • N-terminal amino acid residue A small protein present in all eukaryotic cells tagging protein for destruction ubiquitin
Three enzymes participate in the conjugation of ubiquitin to proteins • Terminal carboxyl of ubiquitin link to a sulfhydril group of E1 • Activated ubiquitin then shuttled to a sulfhydril of E2 • Target protein is tagged by ubiquitin for degradation • Ubiquitin-specific protease recognize the target degrade
2. Oxidation of amino acid residues • Conditions that generate oxygen radicals cause many proteins to undergo mixed-function oxidation of particular residues • Conditions require Fe2+ and hydroxyl radical, and the amino acids most susceptible to oxidation are lysine, arginine, and proline. • E. coli and rat liver contain protease cleaves oxidized glutamine synthetase in vitro, but does not attack the native enzyme • accumulation of oxidatively damaged protein beyond the cell’s capacity to degrade and replace them contribute to importantly to cellular aging
3. PEST sequence all short-lived proteins (i.e., half-lives < 2 h) contain 1 or more regions rich in proline, glutamate, serine, and threonine 4. N-terminal amino acid residue An N-terminal protein residue of Phe, Leu, Tyr, Trp, Lys, or Arg short metabolic lifetimes . Proteins with other termini are far longer-lived. Thus, the intracellular half-life of a particular protein depends on the identity of its N-terminal amino acid residue.