The Intricacies of Signal Transduction in Multicellular Organisms
Signal transduction is essential for the coordination of biochemical reactions in multicellular organisms. It involves intercellular communication where cells send chemical messengers, utilize gap junctions, or interact through cell surface proteins. This process ensures that a signal produced in one cell can influence the behavior of target cells, leading to physiological responses. Key components include receptors, protein kinases, and second messengers, all working in a regulated manner to facilitate effective communication and response to stimuli. Understanding this complex network is crucial for appreciating cellular dynamics and organismal development.

The Intricacies of Signal Transduction in Multicellular Organisms
E N D
Presentation Transcript
Signal transduction ⇌ Whenever I've had to resort to adrenaline to the heart it has never worked, Hema said to herself. Not once. Maybe I do it as a way to signal to myself that the patient is dead. But surely it must have worked, somewhere, with someone. Why else was it taught to us? Cutting for stone. Abraham verghese
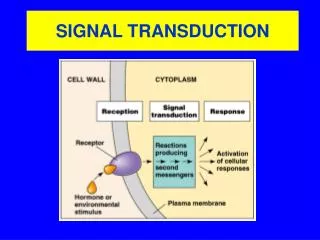
The Structure and Function of Signal Pathways • The enormous structural variety and functional capacity of multicellular organisms is due to their ability to coordinate the biochemical reactions of the various cells of the total organism • The basis for this coordination is the intercellular communication, which allows a single cell to influence the behavior of other cells in a specific manner • Cells can communicate in different ways: • Chemical Messengers: Cells send out signals in the form of specific chemical messengers that the target cell converts into a biochemical reaction • Gap Junctions: Communication between bordering cells is possible via direct contact in the form of “gap junctions” • Cell-cell interaction via cell surface proteins: A cell surface protein of one cell binds a specific complementary protein (or carbohydrate chain) on another cell
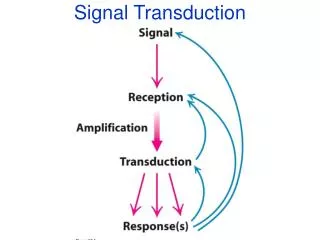
A further intercellular communication mechanism relies on electrical processes. The conduction of electrical impulses by nerve cells is based on changes in the membrane potential • Signal transduction is the process by which cells of a particular type receive and transform a biochemical signal into a physiological reaction • Signaling pathways are involved in the coordination of metabolite flux, the regulation of cell division, differentiation and development of an organism, processing of sensory information Principal Mechanisms of Intercellular Communication
In the communication between cells of an organism, the signals (chemical messengers or electrical signals) are produced in specialized cells • The signal-producing function of these cells is itself regulated, so that the signal is only produced upon a particular stimulus • The following steps are involved in intercellular communication: • Formation of a signal in the signal-producing cell as a result of an external trigger • Transport of the signal to the target cell • Registration of the signal in the target cell • Further transmission of the signal into the target cell • Transformation of the signal into a biochemical or electrical reaction in the target cell • Termination of the signal • Specialized proteins, termed receptors, are utilized for the reception of signals
There are two principal ways by which target cells can process incoming signals: • Cell surface receptors receive the signal at the outside of the cell, become activated and initiate a signaling chain in the interior of the cell. • In such signaling pathways the membrane-bound receptor transduces the signal at the cell membrane so that it is not necessary for the signal to actually enter the cell The Steps of Intercellular Communication
The messenger enters into the target cell and binds and activates the receptor localized in the cytosol or nucleus • Upon receiving a signal, a receptor becomes activated to transmit the signal further • The activated receptor passes the signal onto components, usually proteins, further downstream in the signaling pathway, which then become activated themselves for further signal transmission • A chain of serially operating, intracellular signal transduction processes results • Finally, a specific biochemical process is triggered in the cell, which represents the endpoint of the signaling pathway • Cells possess multiple mechanisms to regulate the intercellular communication as well as the intracellular signal transduction • This allows a specific termination of communication between cells. Often feedback mechanisms are used to adapt the cellular response to the needs of the organism
The key components of intracellular signal transduction include receptors, protein kinases and protein phosphatases, regulatory GTPases, adaptor proteins and second messengers • The regulatory GTPases function as switches that can exist in an active or inactive form • Adaptor proteins mediate the signal transmission between proteins of a signaling chain by bringing these proteins together • They function as clamps to co-localize proteins for an effective and specific signaling • Furthermore, adaptor proteins help to target signaling proteins to specific sub-cellular sites • Second messengers are chemical signaling substances produced or released due to the intracellular activation of enzymes in a signaling chain • Extracellular signaling molecules are released either by exocytosis or passive diffusion into the extracellular space
In special cases, membrane-bound proteins are also used as signaling molecules • Signaling molecules for the communication between cells are known as hormones • Hormones that are proteins and regulate cell proliferation are known as growth factors • The chemical nature of hormones is extremely variable • Hormones can be proteins, peptides, amino acids and amino acid derivatives, derivatives of fatty acids, nucleotides, steroids, retinoids, small inorganic molecules, such as NO • The modification of hormones can lead to compounds that are known as agonists or antagonists • Antagonists are hormone derivatives that bind to a receptor but do not initiate signal transduction • Antagonists block the receptor and thus terminate signal transduction. They have broad medical application since they specifically interfere with certain signal transduction pathways
Various forms of intercellular communication can be discerned based on the range of the signal transmission • In endocrine signaling, the hormone is synthesized in specific signaling, or endocrine cells and exported via exocytosis into the extracellular medium (e.g., blood or lymphatic fluid) • The hormone is then distributed throughout the entire body via the circulatory system so that remote regions of an organism can be reached • Paracrine signal transduction occurs over medium range. • The producing cell must be found in the vicinity of the receiving cells for this type of communication • The signaling is local, and the participating signaling molecules are termed tissue hormones or local mediators • A special case of paracrine signal transduction is synaptic neurotransmission in which a nerve cell communicates with either another nerve cell or with a muscle cell
In autocrine signaling, cells of the same type communicate with one another • If an autocrine hormone is secreted simultaneously by many cells then a strong response occurs in the cells • Autocrine mechanisms are of particular importance in the immune response • A special case of signal transduction is represented by a class of small, reactive signaling molecules, such as NO • NO is synthesized in a cell in response to an external signal and is delivered to the extracellular fluid • Either by diffusion or in a protein-bound form, the NO reaches neighboring cells, and modification of target enzymes ensues, resulting in a change in the activity of these enzymes • NO is characterized as a mediator that lacks a receptor in the classical sense • Signal pathways commonly amplify the initial signal received by the receptor during the course of the signal transduction
SIGNALING BY NUCLEAR RECEPTORS • Nuclear receptors regulate gene expression in response to binding lipophilicmolecules and are thereby involved in the control of a diversity of cellular processes • These proteins are ligand-activated transcription factors that are localized in the cytoplasm and/or in the nucleus • The ligands pass the cell membrane by simple diffusion and bind to the receptors Signal Amplification
By binding to DNA elements in the control regions of target genes the ligand-bound receptor influences the transcription of these genes and thus transmits hormonal signals into a change of gene expression • The naturally occurring ligands of nuclear receptors are lipophilic hormones, among which the steroid hormones, the thyroid hormone T3, and derivatives of vitamin A and D have long been known as central regulators • In recent years it has been recognized that intracellularly formedlipophilic metabolites can also serve as ligands for nuclear receptors and can regulate gene expression through their binding to nuclear receptors • These compounds include prostaglandins and leukotrienes • They also include other molecules synthesized intracellularly as normal metabolites such as fatty acids and bile acids and substances derived from foreign lipophilic substances like drugs
The receptors such as PPAR are quite promiscuous with respect to the nature of the ligand and can bind a broad range of lipophilic ligands • This type of receptors is thought to be involved in the regulation of metabolism and in detoxification • In comparison to signaling pathways which utilize transmembrane receptors , signaling via nuclear receptors is of relatively simple structure • The pathways lead directly, with only a few participating protein components, from the extracellular space to the level of transcription in the nucleus • Nuclear receptors have got separate ligand-binding and DNA-binding domains • That part of the DNA bound specifically by nuclear receptors is known as hormone response element (HRE)
Based on the receptor activation mechanism, the nuclear receptors may be divided into two basic groups • In the first group (those including most of the steroid hormone receptors), the receptors can be localized in the nucleus or in the cytoplasm • The receptors of the other group are always localized in the nucleus. Representative ligands of these receptors are the derivatives of retinoic acid, the T3 hormone and Vit D • The transport of steroid hormones occurs in the form of a complex with a specific binding protein • An example of such a binding protein is transcortin, which is responsible for the transport of the corticosteroids • The steroid hormones enter the cell by diffusion and activate the cytosolic receptors. In the absence of steroid hormones, the receptors remain in an inactive complex- aporeceptor complex • In the aporeceptor complex the receptor is bound to chaperones
The binding of the hormone to the aporeceptor complex leads to activation of the receptor and initiates the translocation of the receptor into the nucleus where it binds its HRE Signal Transduction by Steroid Hormone Receptors
In contrast to signal transduction by the steroid hormone receptors, there are multiple pathways by which the ligands of retinoic acid group are made available for receptor activation • They can follow the classical endocrinological pathway (like vit D), be synthesized inside the cell from inactive precursors (like retinoic acid) or their full synthesis could take place inside a cell (like prostaglandin J2) • In addition, the receptors in this group are found bound with the corresponding HRE in the absence of hormone, acting as repressors of gene activity • In the presence of the hormone an activation of gene expression is usually observed • In general, HREs are composed primarily of two copies of a hexamer DNA sequence • The hexamers can be inverted (palindromic), everted or direct repeats
Inverted Everted Direct Signal Transduction by RXR Heterodimers
The receptors bind to the cognate HRE mainly as dimers, allowing the formation of homodimers as well as heterodimers between various receptor monomers • Very few nuclear receptors are known whose HRE contains only a single copy of the recognition sequence. These receptors bind as monomers to the HRE • The HREs of the steroid hormone receptors possess a palindromic structure; homodimers of receptors are formed • The HRE of the nuclear receptors for all-trans retinoic acid, 9-cis retinoic acid, the T3 hormone and vitamin D usually exhibit a direct repeat of the recognition sequence, resulting in the formation of heterodimers on the DNA • One of the partners in the heterodimer is always the receptor for 9-cis retinoic acid, RXR
SIGNAL TRANSMISSION THROUGH TRANSMEMBRANE RECEPTORS • Another way of transducing signals into the interior of the cell is through transmembrane receptors • Transmembrane receptors are integral membrane proteins, i.e., they possess a structural portion that spans the membrane • An extracellular domain, a transmembrane domain and an intracellular or cytosolic domain can be differentiated within the structure
In many receptors, the extracellular domain contains the ligand-binding site • The function of the transmembrane domain is to pass the signal on to the cytosolic domain of the receptor • Two basic mechanisms are used for conduction of the signal to the cytosol: • Specific protein-protein interactions–the next protein component in the signal transmission pathway, the effector protein, is activated • The conformational change that accompanies the perception of the signal by the receptor creates a new interaction surface for proteins that are located downstream of the receptor • Activation of Enzymes –the arrival of the signal triggers enzyme activity in the cytosolic domain of the receptor, which, in turn, pulls other reactions along with it • The enzyme usually has Tyrosine kinase activity
However, there are other examples where tyrosine phosphatase or Ser/Thr-specific protein kinase activity is activated • The enzyme activity may be an integral part of the receptor, or it may also be a separate enzyme associated with the receptor on the inner side of the membrane • G-Protein Coupled Receptors (GPCR) • Of the transmembrane receptors, the G protein-coupled receptors form the largest single family • GPCR can be activated by extracellular ligands or sensory signals • Extracellular ligands include biogenic amines, such as adrenaline and noradrenaline, histamine, serotonin, lipid derivatives, nucleotides, retinal derivatives, peptides such as bradykinin and large glycoproteins such as luteinizing hormone, and parathormone
Physical stimuli such as light signals are registered and converted into intracellular signals by GPCR; they are also involved in perception of taste and smell • A characteristic structural feature of the GPCR is the presence of 7-transmembrane helices • For the vast majority of 7-helix transmembrane receptors the next downstream located signaling protein is a heterotrimeric GTP/GDP binding protein (G-protein)
When a ligand binds to a GPCR, the structure of the transmembrane part is altered and this change is passed on to the cytoplasmic loops of the receptor • As a consequence, a high-affinity surface is created for binding of the G protein • The G protein, which exists as the inactive GDP form, now binds to the activated receptor and is itself activated • An exchange of GDP for GTP takes place, and the βγ-subunit of the G protein dissociates • Once the G protein is activated, it frees itself from the complex with the receptor, which either returns to its inactive ground state or activates further G proteins • A phenomenon often seen in transmembrane receptors in general, and in G protein-coupled receptors in particular, is desensitization • Desensitization means a weakening of the signal transmission under conditions of long-lasting stimulation
Despite the persistent effect of extracellular stimuli, the signal is no longer passed into the cell interior, or only in a weakened form, during desensitizing conditions • A common way of desensitization is the phosphorylation of the receptor at the cytoplasmic side by specific protein kinases • Phosphorylation of the receptor can be carried out by protein kinase A or protein kinase C • This is a feedback mechanism since PKA and PKC are activated by GPCR • Another way of phosphorylation is through G protein coupled receptor protein kinases (GRK) • The phosphate residues introduced by GRK serve as attachment sites for arrestinwhich serves as a trigger for internalization of the receptor to endosomes
Desensitization of GPCR
The superfamily of GTPasesincludes the heterotrimeric G proteins, the Ras family of small GTPases and the family of initiation and elongation factors • The defining feature of this group is that its members have got a “switch function” • The binding of GTP brings about the transition into the active form (turned on) • Hydrolysis of the bound GTP by the intrinsic GTPase activity converts the protein into the inactive, GDP-bound form (turned off) • Thus, the GTPase activity is one way of terminating signaling • In the case of heterotrimeric G-proteins, the α-subunit has a binding site for GTP or GDP and carries the GTPase activity • Based on comparison of the amino acid sequences, the heterotrimeric G-proteins are divided into four families • The members of the Gs subfamily are activated by hormone receptors, by odor receptors and by taste receptors
Examples include signal transmission by type β-adrenaline receptors and by glucagon receptors • During perception of smell, the smell receptors are activated, and these then pass the signal on via the olfactory G protein Golf • Perception of sweet taste is also mediated via a Gs protein • Transmission of the signal further involves an adenylyl cyclase in all cases, the activity of which is stimulated by the Gs-proteins • The first members of the Gisubfamily to be discovered displayed an inhibitory effect on adenylyl cyclase, hence the name Gi, for inhibitory G proteins • Other members of the Gisubfamily have phospholipase C as the corresponding effector molecule • Type α-2 adrenergic receptors fall into this group • Signal transmission in the vision process is mediated via G proteins known as transducins (Gt)
Perception of bitter taste also takes place via Gi • The Gq subfamily includes G-proteins associated with type α-1 adrenergic receptors • There is also a fourth subfamily known as G12 • Two bacterial toxins, namely pertussis toxin and cholera toxin, were of great importance in determining the function of G- proteins • Both toxins catalyze ADP ribosylation of proteins. During ADP ribosylation, an ADP-ribose residue is transferred from NAD+ to an amino acid residue of a substrate protein • Cholera toxin is an enterotoxin made up of one A subunit (composed of one A1 and one A2 peptide joined by a disulfide link) and five B subunits and has a molecular mass of approximately 84 kDa • In the small intestine, the toxin attaches by means of the B subunits binding to the ganglioside GM1 present in the plasma membrane of mucosal cells
The A subunit then dissociates, and the A1 peptide passes across to the inner aspect of the plasma membrane • It then catalyzes the ADP-ribosylation of the GTP-binding regulatory component (Gs) of adenylyl cyclase, upregulating the activity of this enzyme • Thus, adenylyl cyclase becomes chronically activated resulting in an elevation of cAMP ADP-Ribosylation of of G-Proteins
PKA then phosphorylates the regulatory domains of the cystic fibrosis transmembrane conductance regulator (CFTR) andthe Na+-H+ exchanger • This leads to the inhibition of Na+ absorption and the enhancement of the secretion of Cl- • Thus, massive amounts of NaCl accumulate inside the lumen of the intestine, attracting water by osmosis and contributing to the liquid stools characteristic of cholera • Pertussis toxin is a protein secreted by the bacterium Bordetellapertussis which causes whooping cough • Pertussis toxin carries out an ADP-ribosylation at a cysteine residue of a Gi protein that inhibits adenylyl cyclase, closes Ca2+ channels, and opens K+ channels • The effect of this modification, however, is to lower the G protein's affinity for GTP, effectively trapping it in the "off" conformation. The pulmonary symptoms have not yet been traced to a particular target of the Giprotein
Mechanism of Action of Cholera Toxin
Effector Molecules of G-Proteins • Activated G proteins pass the signal on to subsequent effector molecules that have enzyme activity or function as ion channels • Important effector molecules are adenylyl cyclase, phospolipases, and cGMP-specific phosphodiesterases • The activation of these enzymes leads to concentration changes of diffusible signal molecules such as cAMP, cGMP, diacylglycerol or inositoltriphosphate (IP3), and Ca2+ ,which trigger further specific reactions • cAMP • 3’-5’-cyclic AMP is a central intracellular second messenger that influences many cellular functions, such as gluconeogenesis, glycolysis, lipogenesis, muscle contraction, membrane secretion, learning processes, ion transport, differentiation,…
Concentration of cAMP is controlled primarily by two means, namely via new synthesis by adenylyl cyclase and degradation by phosphodiesterases • Cyclic AMP binds to and activates different signaling proteins • It can regulate ion passage through cAMP-gated ion channels • The majority of the biological effects of cAMP are mediated by the activation of protein kinases. Protein kinases regulated by cAMP are classified as protein kinase A • In the absence of cAMP, protein kinase A exists as a tetramer, composed of two regulatory (R) and two catalytic (C) subunits • In the tetrameric R2C2 form, protein kinase A is inactive since the catalytic center of the C subunit is blocked by the R subunit • Upon binding of four molecules of cAMP, the enzyme dissociates into an R subunit dimer with four molecules of cAMP bound and two free C subunits which are now released from inhibition by the regulatory subunits and can thus phosphorylate Ser/Thr residues on specific substrate proteins
Cyclic-GMP • Like cAMP, 3’-5’-cGMP is a widespread second messsenger • Analogous to cAMP, cGMP is formed by catalysis via guanylyl cyclase from GTP • While adenylyl cyclase is an integral membrane protein, guanylyl cyclase can be found either associated with membranes or as a soluble cytosolic form Cyclic-AMP and Activation of PKA
*cAMP and Gene Transcription • Whereas some responses mediated by cyclic AMP occur within seconds and do not depend on changes in gene transcription, others do depend on changes in the transcription of specific genes • When PKA is activated by cAMP, it enters into the nucleus and phosphorylates a specific gene regulatory protein called cyclic AMP response element-binding (CREB) protein • Phosphorylated CREB then recruits a transcriptional coactivator called CREB-binding protein (CBP) • The CREB/CBP complex binds to CRE on specific genes and activates transcription • This signaling pathway controls many processes in cells, ranging from hormone synthesis (e.g. somatostatin) in endocrine cells to the production of proteins required for the induction of long-term memory in the brain
*Signaling through the βγ –subunit • In some other cases, G proteins directly activate or inactivate ion channels in the plasma membrane of the target cell • Acetylcholine reduces both the rate and strength of heart muscle cell contraction • This effect is mediated by a special class of acetylcholine receptors that activate the Gi protein • Once activated, the α subunit of Gi inhibits adenylyl cyclase while the βγ subunits bind to K+ channels in the heart muscle cell plasma membrane and open them • The opening of these K+ channels makes it harder to depolarize the cell and thereby contributes to the inhibitory effect of acetylcholine on the heart • These acetylcholine receptors, which can be activated by the fungal alkaloid muscarine, are called muscarinic acetylcholine receptors to distinguish them from nicotinic acetylcholine receptors
Nicotinic receptors are ion-channel-coupled receptors on skeletal muscle and nerve cells that can be activated by the binding of nicotine, as well as by acetylcholine
Isozymes of Guanylyl Cyclase • The second messenger function of cGMP is directed towards three targets: cGMP-dependent protein kinases (protein kinase G, PKG), ion channels and cAMPphosphodiesterases • Cyclic GMP carries different messages in different tissues. In the kidney and intestine it triggers changes in ion transport and water retention; in cardiac muscle, it signals relaxation; in the brain it may be involved both in development and in adult brain function
Guanylyl cyclase in the kidney is activated by the hormone atrialnatriuretic factor (ANF), which is released by cells in the atrium of the heart when the heart is stretched by increased blood volume • Carried in the blood to the kidney, ANF activates guanylyl cyclase in cells of the collecting ducts • The resulting rise in [cGMP] triggers increased renal excretion of Na+ and, consequently, of water, driven by the change in osmotic pressure • Water loss reduces the blood volume, countering the stimulus that initially led to ANF secretion • Vascular smooth muscle also has an ANF receptor—guanylyl cyclase; on binding to this receptor, ANF causes vasodilation, which increases blood flow while decreasing blood pressure
A similar receptor guanylyl cyclase in the plasma membrane of intestinal epithelial cells is activated by an intestinal peptide, guanylin, which regulates Cl- secretion in the intestine • This receptor is also the target of a heat-stable peptide endotoxin produced by E. coli and other gram-negative bacteria • The elevation in [cGMP] caused by the endotoxin increases Cl- secretion and consequently decreases reabsorption of water by the intestinal epithelium, producing diarrhea • The soluble guanylylcyclases are regulated by the second messenger NO • They have a heme group that confers NO-sensitivity. NO binding to the heme group results in activation of the guanylyl cyclase activity • Acetylcholine is a vasodilator that acts by causing relaxation of the smooth muscle of blood vessels • However, it does not act directly on smooth muscle
If endothelial cells are stripped away from underlying smooth muscle cells, acetylcholine no longer exerts its vasodilator effect • This indicates that vasodilators such as acetylcholine initially interact with the endothelial cells of small blood vessels via receptors • The receptors are coupled to the phosphoinositide cycle, leading to the intracellular release of Ca2+ through the action of inositoltrisphosphate • In turn, the elevation of Ca2+ leads to the liberation of NO also known as endothelium-derived relaxing factor (EDRF), which diffuses into the adjacent smooth muscle • This leads to the elevation of intracellular levels of cGMP which in turn stimulates the activities of certain PKG, which probably phosphorylate specific muscle proteins, causing relaxation; however, the details are still being clarified
In the heart, cGMP reduces the forcefulness of contractions by stimulating the ion pump(s) that expel Ca+2 from the cytosol • This NO-induced relaxation of cardiac muscle is the same response brought about by nitroglycerin tablets and other nitrovasodilators taken to relieve angina, the pain caused by contraction of a heart deprived of O2 because of blocked coronary arteries • Another important cardiovascular effect of NO is that by increasing synthesis of cGMP, it acts as an inhibitor of platelet aggregation • NO is unstable and its action is brief; within seconds of its formation, it undergoes oxidation to nitrite, nitrate or peroxynitrite • Nitrovasodilators produce long-lasting relaxation of cardiac muscle because they break down over several hours, yielding a steady stream of NO
The effects of increased cGMP synthesis diminish after the stimulus ceases, because a specific phosphodiesterase(cGMP PDE) converts cGMP to the inactive 5-GMP • Humans have several isoforms of cGMP PDE, with different tissue distributions • The isoform in the blood vessels of the penis is inhibited by the drug sildenafil citrate (Viagra), which therefore causes cGMP levels to remain elevated once raised by an appropriate stimulus, accounting for the usefulness of this drug in the treatment of erectile dysfunction • NO is inhibited by hemoglobin and other heme proteins, which bind it tightly • Administration of NO synthase inhibitors to animals and humans leads to vasoconstriction and a marked elevation of blood pressure, indicating that NO is of major importance in the maintenance of blood pressure in vivo
Inositol phospholipids and inositol phosphates • Inositol-containing phospholipids of the plasma membrane are the starting compounds for the formation of various inositol messengers in response to various signals • These messengers include the central second messengers diacylglycerol (DAG) and inositoltrisphosphate (IP3) as well as membrane-bound phosphatidylinositol phosphates (e.g. PIP3) • Phosphatidylinositol is first phosphorylated by specific kinases at the 4’ and 5’ positions of the inositol residue, leading to the formation of phosphatidyl inositol-4,5-bisphosphate (PIP2) • From PIP2, two paths lead to physiologically important messenger substances • One path is phosphorylation to yield PIP3 , which functions as a membrane-localized messenger • The other option is cleavage by phospholipase C, forming the second messengers DAG and IP3
IP3 activates the release of Ca2+ , while DAG acts primarily by stimulation of protein kinase C (PKC) • Phospholipase C can be activated by G-proteins or by transmembrane receptors with intrinsic or associated enzymatic activity • Ca2+ is a ubiquitous signaling molecule whose signaling function is activated by its release from intracellular stores or through Ca2+ entry channels from the extracellular side • The concentration of free Ca2+ in the cytosol of resting cells is very low, about 10–7 M • One reason that the cell tries to keep the free Ca2+ concentration low is the ability of these ions to form poorly soluble complexes with inorganic phosphate • The low concentration of free cytosolic Ca2+ is opposed by a large storage capacity for Ca2+ in specific organelles and vesicles and by a high concentration in the extracellular region where Ca2+ is present at millimolar concentration