Download

1 / 104
1.07k likes | 1.26k Vues
No man is an Iland , intire of it selfe ; every man is a peece of the Continent, a part of the maine ; … any mans death diminishes me, because I am involved in Mankinde ; And therefore never send to know for whom the bell tolls; It tolls for thee. John Donne; Meditation XVII.

E N D
No man is an Iland, intire of it selfe; every man is a peece of the Continent, a part of the maine; … any mans death diminishes me, because I am involved in Mankinde; And therefore never send to know for whom the bell tolls; It tolls for thee. John Donne; Meditation XVII
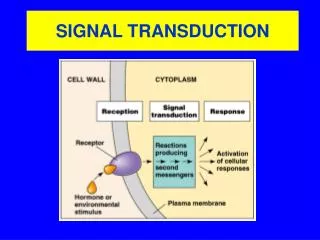
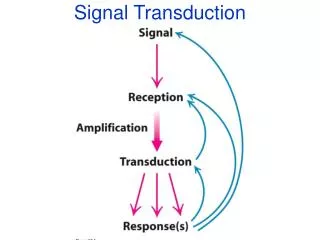
SIGNAL TRANSDUCTION • Even though the efficiency of the tissues is raised through specialization, there should be coordination of activities • Cells communicate and adapt to changes to their environment using various signals • Extracellular signaling molecules or ligands are synthesized and released by signaling cells to produce a specific response only in target cells that have receptors for the signaling molecules • The signal represents information that is detected by specific receptors and converted to a cellular response, which always involves a chemical process • This conversion of information into a chemical change is known as signal transduction
The general pattern of cellular communication Synthesis of signaling molecule Release Transport Binding of the signaling molecule by a receptor protein leading to its activation Initiation of cellular signal transduction pathways by the activated receptor Specific changes in cellular function, metabolism or development Removal of the signal and termination of the cellular response
The majority of receptorsare activated by binding of secreted or membrane-bound molecules • Some receptors, however, are activated by changes in the concentration of a metabolite (e.g., oxygen or nutrients) or by physical stimuli (e.g., light, touch, heat) • based on the distance over which the signal acts, signaling by soluble extracellular molecules can be classified into three types: endocrine, paracrine or autocrine • In endocrine signaling, classic hormones(from the Greek for “set in motion”) travel through the blood and other extracellular fluids to act on target cells far from their site of synthesis • Inparacrine signaling, signaling molecules affect cells in close proximity • neurotransmitters and local hormones
Autocrine signaling refers to the situation in which cells respond to substances that they themselves release • some growth factors, PAF, secretions of tumor cells • Some signaling molecules can be short or long range mediators e.g. epinephrine • Integral membrane proteins can also serve in signaling: • bind with receptors on adjacent cells • cleavage of the exoplasmic portion of the proteins and the fragment acts as a soluble signaling molecule • Features of signal transduction • Specificity – signaling molecule and receptor are complementary • through non-covalent interactions • even when there are receptors, the subsequent molecules may be absent
Sensitivity –very high affinity between ligand and receptor • ligand-receptor interaction may show cooperativity in some cases • the original signal brought by the primary messenger is amplified by enzyme cascades: the number of activated mediators shows a progressive increase • the constant presence of a signal leads to the desensitization of receptors; sensitivity is restored when the level of the stimulus falls below a certain threshold • there may also be upregulation or downregulation of receptors on the surface of the cell • Integration – the ability of the system to receive multiple signals and produce a unified response appropriate to the needs of the cell or organism • Signaling pathways interact with each other at different levels
In terms of their precursors , signaling molecules fall into six groups: • peptides – the largest group; comprising of most classic hormones, growth factors and cytokines • vary from small peptides (TRH = 3 amino acids) to big ones (PTH = 84 a a) • steroids –hormones of the adrenal cortex, and the gonads • amino acids and their derivatives – thyroid hormones and several neurotransmitters • fatty acid derivatives –eicosanoids, PAF • vitamins– vitamins A and D • nucleotide derivatives – adenosine, ADP, ATP
The synthesis of peptide signaling molecules • Like all proteins, they are synthesized by DNA transcription and mRNA translation • The primary transcript of the hormone may code for an inactive prohormone which undergoes proteolysis to give the active hormone • e.g. preproinsulin→proinsulin→insulin; the prohormone of ACTH is cleaved to active ACTH, β-endorphin and melanocyte stimulating hormone (MSH) • Some peptide hormones undergo glycosylation in the Golgi • The individual components of dimeric peptide hormones (TSH, hCG, LH, FSH) are encoded by separate genes • they share a common α subunit but possess specific β subunits • Steroid hormones also may need activation; e.g. testosterone has to be reduced to 5αdihydrotestosterone
Hormone transport • Peptides are hydrophilic and travel in the blood unbound • Steroid hormonesand T4 are complexed with specific binding proteins like cortisol binding globulin (CBG), sex hormone binding globulin (SHBG),… • Albumin may serve in non-specific steroid hormone transport • Hormone inactivation • Inactivation of molecules may be specific (enzyme-based) or non-specific mechanisms • Steroid hormones are biotransformed in the liver and removed through the urine • The products of MAO and COMT action on catecholamines and other neurotransmitters are removed through the urine as they are or may be further modified by the liver before excretion
Peptide hormones are completely degraded and the amount of active peptide hormones present in the urine is negligible • Eicosanoids are inactivated through the chemical modification of groups essential for their activity and they ultimately give water-soluble products like dicarboxylic acids • The hierarchy of neuronal and hormonal signals • Most of the classical endocrine glands are functionally hierarchical; a notable exception being pancreatic islets • A hormone secreted from one gland activates another gland to produce another hormone, which in turn activates the target tissue –an axis is formed • Such axes constitute an ‘amplification cascade’ (even before reaching the level of enzymatic cascades) allowing the primary signal to be enhanced several fold • Negative feedback regulation at different levels; positive feed back rare e.g. increasing production of oxytocin by the pituitary in response to cervical stretch during childbirth
Categories of signaling mechanisms • There are six basic types of signaling mechanisms: • gated ion channels • receptor enzymes • receptors that activate enzymes through G-proteins • receptors that activate enzymes without the involvement of G-proteins • intracellular receptors • adhesion receptors that recognize signaling molecules bound with cells • Please read a bit about ligand-gated and voltage-gated channels; the role of integrins in the extracellular matrix
RECEPTOR ENZYMES • They usually have a ligand-binding extracellular domain and a cytosolic domain with enzymatic activity • The two domains are connected by a single transmembrane domain • The enzyme can have tyrosine kinase, serine/threonine kinase or guanylylcyclase activities • Tyrosine kinases • The insulin receptor • Is an α2β2 dimer bound by disulfide bridges • The insulin binding site is on the αsubunits • The enzymatic activity is in the cytosolic part of the βsubunits • First there is auto(cross)phosphorylation of tyrosine residues in the C-termini of the subunits • Then comes the phosphorylation of other proteins
The main protein phosphorylated by the insulin receptor is called insulin receptor substrate 1 (IRS-1) • The phosphorylated IRS-1 acts like a ‘docking’ site for a system of adaptor proteins which conduct the signal further • The signal maybe destined for the cytosol or the nucleus • The cytosol • Phosphoinositide 3-kinase (PI-3k) binds IRS-1 • the phosphotyrosine residues on IRS-1 are recognized by an SH2 (Src homology 2) domain on PI-3K • PI-3K phosphorylatesphosphatidylinositol 4,5-bisphosphate (PIP2) to phosphatidylinositol 3,4,5-bisphosphate (PIP3) • PIP3 binds protein kinase B (PKB); the binding favors the phosphorylation and activation of PKB by PDK 1 • PKB phosphorylates Ser and Thr residues on target proteins like glycogen synthase kinase (GSK ) 3, and Glut 4
PKB is also involved in the pathway activated by Δ9-tetrahydrocannabinol (THC) • THC binds to the receptors of endocannabinoids (such as anandamide ) which is synthesized from arachidonate and phosphatidylethanolamine • In addition to the phosphorylation of proteins, insulin exerts many of its effects through dephosphorylation • Dephosphorylation activates glycogen synthase and pyruvate dehydrogenase but it inactivates glycogen phosphorylase and hormone-sensitive lipase • It is unclear how insulin activates phosphoproteinphosphatases THC
the PKB signal can be terminated by a phosphatase that removes the phosphate at the 3 position of PIP3 • The nucleus • Insulin exerts its effects on genetic expression (of enzymes that control metabolism and the cell cycle) through a different pathway • The SH2 domain of a protein known as Grb 2 attaches to phosphotyrosine residues on IRS-1 • Through its SH3 domain Grb 2 attaches to Sos • Sos catalyzes the replacement of GDP for GTP on Ras • 3 kinases – Raf-1, MEK and ERK – are consecutively activated • ERK enters the nucleus and activates proteins that serve as transcription factors • ERK is a member of a family of enzymes known as mitogenactivated protein kinases (MAPK)
there are many other receptor enzymes which have receptor tyrosine kinase activities • epidermal growth factor (EGF),platelet-derived growth factor (PDGF), nerve growth factor (NGF),… • EGF is a small, soluble peptide hormone that binds to cells in the embryo and in skin and connective tissue in adults, causing them to divide • It is synthesized as an integral plasma membrane protein • Membrane-bound EGF can bind to and signal an adjacent cell by direct contact • Cleavage by an extracellular protease releases a soluble form of EGF, which can signal in either an autocrine or a paracrine manner • PDGF stimulates growth and tissue repair in an injured region • Except the insulin receptor, the other receptors in the RTK group dimerize upon the binding of ligand
Receptor Serine/Threoninekinases • The group includes receptors for members of the transforming growth factor β (TGF β) superfamily • Isoforms of TGFβsuperfamily are inhibitors of proliferation • Loss of TGFβ receptors or certain intracellular signal transduction proteins in the TGFβ pathway, thereby releasing cells from this growth inhibition, frequently occurs in tumors • TGFβ also promotes expression of cell-adhesion molecules and extracellular-matrix molecules • TGFβ signals certain types of cells to synthesize and secrete growth factors that can overcome the normal TGF β -induced growth inhibition; this explains why TGF was originally detected as a growth factor • The TGFβ receptor complex is composed of two different single membrane-spanning receptor subunits (type I and II)
TGFβ binds to a type II receptor and the activated type II receptor recruits a type I receptor which it phosphorylates at a serine residue, forming an activated receptor complex. • The type I receptor then binds a receptor-specific Smad protein (called R-Smad) which it phosphorylates at serine residues • The phosphorylated R-Smad undergoes a conformational change and dissociates from the receptor • R-Smad forms a complex with another member of the Smad family, Smad 4 (Smad 4 is known as the common Smad, Co-Smad, and is not phosphorylated) • The Smad complex, which may contain several Smads, translocates to the nucleus, where it activates or inhibits the transcription of target genes • Receptors for different ligands bind different Smads, which bind to different sites on DNA and regulate the transcription of different genes
Within the nucleus R-Smads are being continuously dephosphorylated, which results in the dissociation of the R-Smad/co-Smad complex and export of these Smads from the nucleus ----- signal termination
Guanylylcyclase activity • Atrialnatriuretic factor (ANF), NO (EDRF), intestinal guanylin • the binding of signaling molecules leads to the synthesis of cGMP from GTP • cGMP activates cGMP-dependent protein kinase also known as protein kinase G (PKG) • PKG phosphorylates various proteins at Ser and Thr residues leading to the physiological effects of the mediators • guanylin increases the secretion of Cl_ in the intestine • the receptor for guanylin can also bind the toxin produced by E. coli and other bacteria • The signaling is terminated when cGMP is converted to GMP by a variety of phosphodiesterases
G-PROTEIN-COUPLED RECEPTORS (GPCR) • The family includes receptors for several hormones and neurotransmitters, light-activated receptors (rhodopsins) in the eyes, odorant and taste receptors • The defining features are: • a plasma membrane receptor with seven membrane- spanning regions (7tm; serpentine receptor) with the N-terminal segment on the exoplasmic face and the C-terminal segment on the cytosolic face • an enzyme in the plasma membrane that generates an intracellular second messenger • a guanosine nucleotide–binding protein (G protein) • G-proteins contain three subunits:α, βandγ • During intracellular signaling the β and γsubunits remain bound together and are usually referred to as the Gβγsubunit
The Gα subunit is a GTPase switch protein that alternates between an active (on) state with bound GTP and an inactive (off) state with bound GDP • Binding of the receptor by the appropriate signaling molecule activates the G- protein and the activated G-protein in its turn activates an associated effector protein • The effector protein may be inhibited in some cases • The effector protein may be: • adenylylcyclase (AC) that converts ATP to cAMP • phospholipase C (PLC) which converts PIP2 to diacylglycerol (DAG) and inositol 1,4,5-triphosphate (IP3) • cAMP, IP3 and DAG are second messengers; so is cGMP • The second messengers activate a range of proteins leading to different physiological effects • Hydrolysis of GTP to GDP (by an intrinsic ATPase) causes Gαto dissociate from the effector and reassociate with the Gβγsubunit
the mechanism of β1 and β2-adrenergic receptors is the prototype for the action of GPCR • Signaling through the Gs protein leads to lipolysis in the adipose, glycogenolysis in the liver and the muscles, contraction of cardiac muscles • In the liver glucagon and epinephrine bind to different receptors, but both receptors interact with and activate the same Gs • The α1-adrenergic receptor is coupled to a Giprotein that inhibits AC • The Gq protein coupled to the α2-adrenergic receptor activates a different effector enzyme that generates different second messengers • cAMP exerts its effects through the activation of cAMP-dependent protein kinase also known as protein kinase A (PKA)
Inactive PKA is a tetramer consisting of two regulatory (R) subunits and two catalytic (C) subunits • Each R subunit has two distinct cAMP-binding sites • binding of cAMP to both sites in an R subunit leads to release of the associated C subunit unmasking its catalytic site and activating its kinase activity • PKA phosphorylates and inactivates glycogen synthase • PKA also phosphorylates glycogen phosphorylase kinase; glycogen phosphorylase kinase phosphorylates glycogen phosphorylase • The net effect is an increase in the amount of free glucose • Phosphoproteinphosphatases (PP) reverse the effect of PKA by removing the phosphates added to different enzymes through the action of PKA • PKA regulates the activity of PP:PKA phosphorylates and activates an inhibitor of PP
Epinephrine levels as low as 10-10 M can stimulate liver glycogenolysis • 10-6 M cAMP can be produced • The amount of glucose released can reach 10-2 M • In skeletal muscles the ratios of the concentrations of PKA:GPK:GP is 1:10:240 • SIGNAL AMPLIFICATION
Just like in the case of cGMP, phosphodiesterase converts cAMP to AMP terminating the signaling • Insulin activates phosphodiesterase • Methylxanthinessuch as caffeine and theophyline inhibit phosphodiesterase; cAMP persists and mobilization of fuels increases • In the continuous presence of epinephrine, the receptors are desensitized • The receptor is phosphorylated both by β-adrenergic receptor kinase (βARK) and PKA • The phosphorylation creates a binding site for the protein β-arrestin (βarr) • The binding of β-arrestin inhibits the interaction of the receptor with G-proteins; it also facilitates the removal of the receptors from the cell membrane by endocytosis
Toxins produced by Vibriocholerae, which causes cholera, and several other enteric bacteria alter the permeability barrier of the intestinal epithelium by altering the composition or activity of tight junction • Thecholera A toxin catalyzes the transfer of ADP-ribose from NAD+ to the subunit of Gs blocking its GTPase activity; Gs will be permanently activated • The continuous activation of AC of intestinal epithelial cells and the subsequently high levels of cAMP trigger constant secretion of Cl-, HCO3- and water into the intestinal lumen • Bordetellapertussis, a bacterium that commonly infects the respiratory tract, is the cause of whooping cough • Pertussis toxin catalyzes a modification of Gi that prevents release of bound GDP, thus locking Gi in the inactive state • This inactivation of Gi leads to an increase in cAMP in epithelial cells of the airways, promoting loss of fluids and electrolytes and mucus secretion
The effect of cholera toxin on intestinal G-proteins
cAMP and genetic expression • Some of the catalytic subunits of PKA translocate to the nucleus and phosphorylate serine-133 on cAMP-response element binding (CREB) protein • Phosphorylated CREB associates with the co-activator CBP/P300 • The CREB/ CBP/P300 complex binds to cAMP-response elements (CRE) on the DNA leading to the transcription of genes for proteins involved in various processes • e.g., the production of somatostatin (a peptide that inhibits the release of various hormones); gluconeogenetic enzymes in the liver • This means that cAMP, in addition to the relatively swift modulation of the activities of various enzymes, also activates a slower response pathway, gene transcription