Download

1 / 69
760 likes | 1.08k Vues
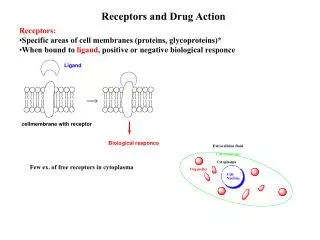
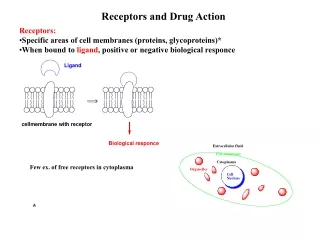
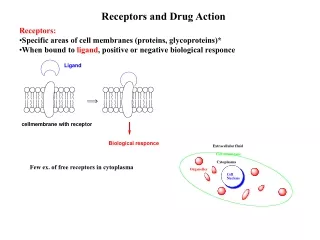
The Organic Chemistry of Drug Design and Drug Action. Chapter 4 Enzymes. Enzymes. Just like drug-receptor complexes, enzymes form complexes with substrates. Two characteristics of enzymes: recognize a substrate catalyze a reaction with it.

E N D
The Organic Chemistry of Drug Design and Drug Action Chapter 4 Enzymes
Enzymes • Just like drug-receptor complexes, enzymes form complexes with substrates. • Two characteristics of enzymes: • recognize a substrate • catalyze a reaction with it
Enzymes - proteins that catalyze reactions in a biological system. They function by lowering transition state energies and by raising ground state energies. Pauling (1946) - Enzyme is a flexible template designed to be complementary to structure of substrate at the transition state of the reaction, not the ground state. As the reaction proceeds toward the transition state, the enzyme interacts more effectively, which accelerates the reaction - called transition state stabilization. Conformational change occurs to align catalytic groups and destabilize the enzyme - induced fit hypothesis.
Similar to noncatalytic receptors, substrate binds to a small part of enzyme - called the active site. • Two key factors of enzyme catalysis: • specificity • rate acceleration Active site contains amino acid side chains used in catalysis. • Some enzymes require cofactors (also called coenzymes): • organic molecules • metal ions
Specificity of Enzyme Reactions specificity of binding specificity of reaction Enzyme catalysis initiated by formation of ES complex (Michaelis complex) Involve same interactions as drug-receptor complex
Scheme 4.1 Michaelis complex Binding specificity can be absolute (only one substrate) or very broad (many substrates). Enzymes are chiral catalysts (all L-amino acids) Interaction with a racemic mixture gives two diastereomeric complexes Therefore, different energies, different reactivities. Both ES complexes may form, but only one may lead to product.
Resolution of Racemic Mixtures with Chiral Reagents Scheme 4.2 Diastereomeric salts
Reaction specificity depends on • Acidic, basic, and nucleophilic groups of the active site amino acids • Coenzymes (also called cofactors)-specific organic molecules or metal ions
Specificity for chemically identical protons Figure 4.2 Only Ha is removed stereospecifically
Catalysts stabilize transition state energy relative to ground state, which decreases G‡ (Ea) Rate Acceleration TS‡ Ea Figure 4.3 Chemical catalyst Enzyme Enzyme stabilizes TS‡, destabilizes ES, destabilizes intermediates.
Mechanisms of Enzyme Catalysis • Most common: • Approximation, • Covalent catalysis, • General acid-base catalysis, • Electrostatic catalysis, • Desolvation, • And strain
Approximation • Rate acceleration by proximity • After the ES complex forms, the reaction becomes 1st order rather than 2nd order. • Equivalent to increasing the concentration of the reacting groups.
Consider a 2nd order reaction Scheme 4.3
Model for Approximation Effective molarity (EM) - concentration of the catalytic group required to cause the intermolecular reaction to proceed at the observed rate of the intramolecular reaction. Table 4.1
Covalent Catalysis Scheme 4.4 Enzymatic analogy to anchimeric assistance (neighboring group assistance) Common active site nucleophiles: - SH (Cys) - OH (Ser) (His) - NH2 (Lys) - COOH (Glu/Asp)
Anchimeric assistance by a neighboring sulfur atom Scheme 4.5
General Acid-Base Catalysis Important whenever proton transfer occurs Two kinds of acid-base catalysis: specific and general Specific acid-base catalysis is determined by [H3O+] or [HO-] (i.e., the pH). General acid-base catalysis occurs by an increase in buffer concentration at a constant pH.
Specific Acid-Base Catalysis Consider poor electrophile poor nucleophile Scheme 4.6
Increase pH Scheme 4.7 excellent nucleophile
Decrease pH Scheme 4.8 excellent electrophile
General Acid-Base Catalysis Unlike reactions in solution, enzymes can use acid and base catalysis simultaneously. Scheme 4.9 general acid general base pKa values of amino acids in the active site are not necessarily the same as in solution
Example of General Acid-Base Catalysis Scheme 4.10 -chymotrypsin
Electrostatic Catalysis oxyanion hole transition state stabilization Scheme 4.11
Desolvation Desolvation (removal of H2O molecules) of charged groups destabilizes the ground state (less polar environment); therefore groups are more reactive. Also, after desolvation, the charged groups in Scheme 4.11 are more able to stabilize charge in the transition state.
Strain or Distortion Scheme 4.12 Because of ring strain, 4.4 is much more reactive than 4.5. Strain induced in the enzyme or in the substrate raises the ground state energy (induced fit theory). Conformational change may be used to induce strain energy.
Destabilization of the ground state is necessary to assure smaller activation energies. Figure 4.4 Rate accelerations of 1010 - 1014 times nonenzymatic reactions. Product release sometimes is slow step.
Scheme 4.13 Example of Enzyme Catalysispeptidoglycan transpeptidase
Coenzymes (or Cofactors) • Organic molecules or metal ions essential for catalytic action • Most organic cofactors are derived from vitamins (essential nutrients). • Only coenzymes whose chemistry will be utilized throughout the remainder of the course are discussed.
Pyridoxal 5’-phosphate (PLP)(from vitamin B6) Catalyzes reactions of amino acids Table 4.4
PLP PLP bound at active site abbreviated structure Figure 4.5
First Step in All PLP-Dependent Reactions Scheme 4.14 From here all of the PLP reactions occur
Pyridinium group can stabilize electrons by resonance from the C-H, C-COO-, or C-R bonds. To get regiospecific cleavage the bond that breaks must lie in a plane perpendicular to the plane of the PLP-imine -e- system. Figure 4.6 C-H bond is perpendicular to the system How do you freeze free rotation and control which bond is perpendicular?
Dunathan Hypothesis (1971) A cationic group could interact with the carboxylate to control the positions of the bonds perpendicular to the -system. Figure 4.7
PLP Racemases Scheme 4.15 All steps are reversible Keq = 1
Decarboxylases Scheme 4.16
Aminotransferases(formerly called transaminases) Scheme 4.17
First Half Reaction of Aminotransferases Scheme 4.18
SecondHalf Reaction of Aminotransferases Scheme 4.19
-Elimination When X is a leaving group, elimination can occur. Scheme 4.20
Tetrahydrofolate and Pyridine Nucleotides Coenzymes that transfer one-carbon units are derived from the vitamin folic acid. folic acid
Reduction of two C=N double bonds to give tetrahydrofolate requires another coenzyme, reduced nicotinamide adenine dinucleotide (NADH). tetrahydrofolate
Reduced nicotinamide adenine dinucleotide coenzymes are derived from vitamin B3 (niacin). part vital to the chemistry When R’ = H NADH When R’ = PO3= NADPH abbreviated structure
NAD(P)H reacts like a hydride reducing agent to reduce the imine bonds. Scheme 4.21 folic acid dihydrofolate tetrahydrofolate Tetrahydrofolate is involved in reactions that transfer one-carbon units from one substrate to another.
Different pyridine nucleotide enzymes use only one of the two hydrogens at C-4. Because the enzyme can differentiate these chemically equivalent hydrogens, they are named differently. An atom is prochiral if by changing one of its substituents, it is converted from achiral to chiral. Prochiral H’s - if HR is changed to 2H, then the carbon becomes chiral (R stereochemistry) pro-S hydrogen pro-R hydrogen
The carbon atom that is transferred is derived from serine in a PLP-dependent -cleavage reaction. atom to be transferred Scheme 4.22
Because of the toxicity of formaldehyde, the reaction in Scheme 4.22 does not occur until the acceptor (tetrahydrofolate) is bound. Scheme 4.23 N10-methylenetetrahydrofolate N5 -methylenetetrahydrofolate N5 ,N10-methylenetetrahydrofolate (Keq = 3.2 x 104) These forms of the coenzyme can transfer a hydroxymethyl group to substrate.
N5 ,N10-Methylenetetrahydrofolate can be oxidized by a NADP+-dependent enzyme to give N5 ,N10-methylenyltetrahydrofolate. Scheme 4.24 hydrolysis gives N10-formyltetrahydrofolate N5-formyltetrahydrofolate
N5 ,N10-Methylenetetrahydrofolate can be reduced by a NADPH enzyme to give N5-methyltetrahydrofolate. Scheme 4.25
Biosynthesis of Purines Example of a one-carbon transfer at the formate oxidation state. Scheme 4.26 inosine MP