Bases and nucleotides
Bases and nucleotides The nucleic acids play a central role in the storage and expression of genetic information.


Bases and nucleotides
E N D
Presentation Transcript
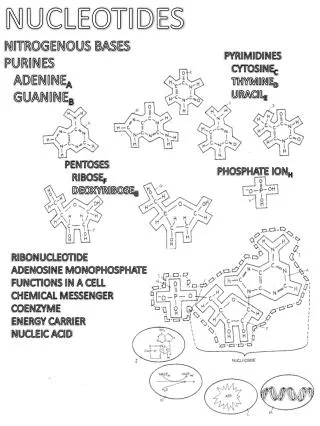
Bases and nucleotides The nucleic acids play a central role in the storage and expression of genetic information. They are divided into two major classes: (1)deoxyribonucleic acid (DNA) functions solely in information storage, (2)ribonucleic acids (RNAs) are involved in most steps of gene expression and protein biosynthesis. All nucleic acids are made up from nucleotide components, consist of a base, a sugar, and a phosphate residue
GULA RIBOSA 5 4 1 3 2 • Gula pada asam nukleat adalah ribosa. • Ribosa (b-D-furanosa) adalah gula pentosa (jumlah karbon 5). • Perhatikan penomoran. • Dalam penulisan diberi tanda prime(') untuk membedakan penomoran pada basa nitrogen KRT-2011
PERHATIKAN • Ikatan gula ribosa dengan basa nitrogen (pada atom karbon nomor 1). • Ikatan gula ribosa dengan gugus fosfat (pada atom karbon nomor 5). • Gugus hidroksil pada atom karbon nomor 2 KRT-2011
BASA NITROGEN • Basa nitrogen berikatandenganikatan-b pada atom karbon nomor1' darigularibosaataudeoksiribosa. • Pirimidinberikatankegularibosapada atom N-1 daristrukturcincinnya. • Purinberikatankegularibosapada atom N-9daristrukturcincinnya. KRT-2011
BASA PIRIMIDIN DAN PURIN Perhatikan struktur cincinnya KRT-2011
GUGUS FOSFAT Nukleosida (Gula Ribosa yang berikatan dengan basa nitrogen) + satu atau lebih gugus fosforil disebut nukleotida. KRT-2011
Nucleic acid bases • The bases that occur in nucleic acids are aromatic heterocyclic compounds derived from either pyrimidine or purine. • The purine bases adenine (abbreviation Ade, not “A”!) and guanine • (Gua) and the pyrimidine base cytosine • (Cyt) are present in both RNA and DNA. In • contrast, uracil (Ura) is only found in RNA. In • DNA, uracil is replaced by thymine (Thy),
When a nucleic acid base is N-glycosidically linked to ribose or 2-deoxyribose, it yields a nucleoside. • The nucleoside adenosine (abbreviation: A) is formed in this way from adenine and ribose, for example. • The corresponding derivatives of the other bases are called guanosine (G), uridine (U), thymidine(T) and cytidine (C). • When the sugar component is 2-deoxyribose, the product is a deoxyribonucleoside—e. g., 2-deoxyadenosine (dA, not shown).
In the cell, the 5OH group of the sugar component of the nucleoside is usually esterified with phosphoric acid. • If the 5phosphate residue is linked via an acid–anhydride bond to additional phosphate residues, it yields nucleoside diphosphates and triphosphates—e. g., ADP and ATP, which are important coenzymes in energy metabolism. All of these nucleoside phosphates are classified as nucleotides.
Polynucleotides consisting of ribonucleotide components are called ribonucleic acid (RNA), while those consisting of deoxyribo-nucleotide monomers are called deoxyribonucleicacid (DNA).
RNA • Ribonucleic acids (RNAs) are polymers consisting of nucleoside phosphate components that are linked by phosphoric acid diester bonds. • The bases the contain are mainly uracil, cytosine, adenine, and guanine, but many unusual and modified bases are also found in RNAs (B). • RNAs do not form extended double helices. In RNAs, the base pairs usually only extend over a few residues. • Large RNAs such as ribosomal 16SrRNA (center) contain numerous “stem and loop” regions of this type. These sections are again folded three-dimensionally—i. e., like proteins, RNAs have a tertiary structure
RNA • Cellular RNAs vary widely in their size, structure, and lifespan. The great majority of them are ribosomal RNA (rRNA), which in several forms is a structural and functional component of ribosomes. • Ribosomal RNA is produced from DNA by transcription in the nucleolus, and it is processed there and assembled with proteins to form ribosome subunits. • The bacterial 16S-rRNA shown in Fig. A, with 1542 nucleotides (nt), is a component of the small ribosomae subunit, while the much smaller 5S-rRNA (118 nt) is located in the large subunit • Messenger RNAs (mRNAs) transfer genetic information from the cell nucleus to the cytoplasm. • The transfer RNAs (tRNAs) function during translation as links between the nucleic acids and proteins
DNA • (DNAs) are polymeric molecules consisting of nucleotide building blocks. • DNA contains 2-deoxyribose, and the uracil base in RNA is replaced by thymine. • DNA consists of two polydeoxynucleotide molecules (“strands”). Each base in one strand is linked to a comple-mentarybase in the other strand by H-bond. (A =T; G= C) • Potential donors are amino groups (Ade, Cyt, Gua) and ring NH groups. Possible acceptors are carbonyl oxygen atoms (Thy, Cyt, Gua) and ring nitrogen atoms. Two linear and therefore highly stable bonds can thus be formed in A–T pairs, and three in G–C pairs • DNA serves to store genetic information. Specific segments of DNA (“genes”) are transcribed as needed into RNAs, which either carry out structural or catalytic tasks themselves or provide the basis for synthesizing proteins
All of the words (“codons”) contain three letters (“triplets”), and each triplet stands for one of the 20 proteinogenic amino acids.
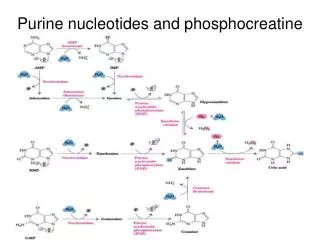
The principles underlying the degradation of purines (1) and pyrimidines (2) differ. • In the human organism, purines are degraded into uric acid and excreted in this form. The purine ring remains intact in this process. • In contrast, the ring of the pyrimidine bases (uracil, thymine, and cytosine) is broken down into small fragments, which can be returned to the metabolism Degradation of nucleotides
Purine (left). • The purine nucleotide guanosinemonophosphate (GMP, 1) is degraded in two steps—first to the guanosine and then to guanine (Gua). Guanine is converted by deaminationinto another purine base, xanthine.
Purine and pyrimidine biosynthesis • The bases occurring in nucleic acids are derivatives of the aromatic heterocyclic compounds purine and pyrimidine • The major intermediates in the biosynthesis of nucleic acid components are the mononucleotides uridinemonophosphate(UMP) in the pyrimidine series and inosinemonophosphate (IMP, base: hypoxanthine) in the purines. • The synthetic pathways for pyrimidines and purines are fundamentally different. For the pyrimidines, the pyrimidine ring is first constructed and then linked to ribose 5-phosphate to form a nucleotide. By contrast, synthesis of the purines starts directly from ribose 5-phosphate.
Purine and pyrimidine biosynthesis • The precursors for the synthesis of the pyrimidinering are carbamoyl phosphate, which arises from glutamate and HCO3 – (1a) and the amino acid aspartate. • These two components are linked to N-carbamoylaspartate(1b) and then converted into dihydroorotateby closure of the ring (1c). In mammals, steps 1a to 1c take place in the cytoplasm, and are catalyzed by a singlemultifunctional enzyme. • In the next step (1d), dihydroorotate is oxidized to orotate by an FMN-dependent dehydrogenase. • Orotate is then linked with phosphoribosyldiphosphate (PRPP) to form the nucleotide orotidine 5-monophosphate (OMP). Finally, decarboxylation yields uridine 5-monophosphate (UMP).
Purine and pyrimidine biosynthesis • Purine biosynthesis starts with PRPP. • Formation of the ring starts with transfer of an amino group, from which the later N-9 is derived (2a). • Glycine and a formyl group from N10-formyl-THF then supply the remaining atoms of the five-membered ring (2b, 2c). • Before the five-memberedring is closed (in step 2f), atoms N-3 and C-6 of the later six-membered ring are attached (2d, 2e). • Synthesis of the ring then continues with N-1 and C-2 (2g, 2i). In the final step (2j), the six-membered ring is closed, and inosine 5-monophosphate arises. • The IMP formed does not accumulate, but is rapidly converted into AMP and GMP.
Nucleotide biosynthesis • De novo synthesis of purines and pyrimidines yields the monophosphates IMP and UMP, respectively. • All other nucleotides and deoxynucleotides are synthesized from these two precursors. • The synthesis of purine nucleotides (1) starts from IMP. The base it contains, hypoxanthine, is converted in two steps each into adenine or guanine. The nucleoside monophosphatesAMP and GMP that are formed are then phosphorylated by nucleoside phosphate kinases to yield the diphosphatesADP and GDP, and these are finally phosphorylated into the triphosphatesATP and GTP. • The nucleoside triphosphates serve as components for RNA, or function as coenzymes (see p. 106). Conversion of the ribonucleotides into deoxyribonucleotides occurs at the level of the diphosphatesand is catalyzed by nucleoside diphosphatereductase (B).
The biosynthetic pathways for the pyrimidine nucleotides (2) are more complicated. • The first product, UMP, is phosphorylated first to the diphosphate and then to the triphosphate, UTP. • CTP synthasethen converts UTP into CTP. • Since pyrimidine nucleotides are also reduced to deoxyribonucleotides at the diphosphate level, CTP first has to be hydrolyzed by a phosphatase to yield CDP before dCDP and dCTP can be produced. • The DNA component deoxythymidinetriphosphate • (dTTP) is synthesized fromUDP in several steps. The base thymine, which only occurs in DNA, is formed by methylation of dUMP at the nucleosidemonophosphate level
Ribonucleotide reduction • 2-Deoxyribose, a component of DNA, is not synthesized as a free sugar, but arises at the diphosphate level by reduction of ribonucleosidediphosphates. • This reduction is a complex process in which several proteins are involved. The reducing equivalents needed come from NADPH+H+. • However, they are not transferred directly from the coenzyme to the substrate, but first pass through a redox series that has several steps (1). • In the first step, thioredoxinreductase reduces a small redox protein, thioredoxin, via enzyme-bound FAD. This involves cleavage of a disulfide bond in thioredoxin. The resulting SH groups in turn reduce a catalytically active disulfide bond in nucleoside diphosphatereductase (“ribonucleotidereductase”). • The free SH groups formed in this way are the actual electron donors for the reduction of ribonucleotidediphosphates.
Hampir semua organisme mampu mensintesis nukleotida dr prekursor yg lebih sederhana jalur de novo untuk nukleotida mirip utk setiap organisme • Nukleotida juga dapat disintesis dari hasil pemecahan nukleotida yang telah ada salvage pathway (recycle) yaitu dari degradasi pirimidin dan purin dari sel yang mati (regenerasi) atau dari makanan
5-Phospho- -D-ribosyl-1-pyrophosphate (PRPP) • Intermediet untuk baik proses de novo and salvage pathway • Berasal dari ribosa 5 phosphat
IMP synthase GAR synthetase AICAR transformylase GAR transformylase SAICAR lyase FGAR amidotransferase SAICAR synthetase FGAM cyclase AIR karboksilase
Hal-hal penting dalam sintesis de novo purin: • Sangat tergantung pada “pool” ribosa • Gugus amina didonor oleh glutamin dgn enzim amidotransferase • Glisin dan fumarat donor ring dlm nukleotida • Daur reaksi dikontrol secara alosterik dgn AMP, ADP, GMP dan GDP. bekerja pada PRPP amidotransferase
Daur diawali dgn perubahan PRPP IMP • IMP = Inosine monofosfat mrpkn bentuk nukleotida purin yang pertama dibentuk dlm daur ini • Sebagai basa adalah hypoxanthin
Adenilosuksinat synthetase IMP dehidrogenase XMP aminase Adenilosuksinat lyase DAUR dr IMP AMP & GMP
CP synthetase Aspartattranscarbomoylase Dihidrooratate DH dihydrorotase Metabolisme de novo nukleotidapirimidine
Orotat fosforibosiltransferase Orotidilate dekarboksilase CTP synthetase UMP kinase Nukleosida diphosphat kinase
Hal-hal penting dalam sintesis de novo pirimidine: • cincin pirimidine disintesis terpisah dr gula ribosa nya • Daur pirimidine de novo tidak bercabang produk akhir dr daur adalah UMP yang mrpkn bahan dari CMP • Reaksi pertama pembtkan karbamoyl aspartate dr asp dan carbomoyl-P titik regulasi yg penting dlm daur tsb • Aspartat transcarbomoylase (ATCase) diaktivasi oleh diaktivasi oleh ATP dan dihambat oleh CTP sbg produk akhir