Download

1 / 22
260 likes | 586 Vues
Lecture 18: Intracellular transport. Flint et al., Chapter 12. General remarks. Viruses are cellular parasites Eukaryotic cells are functionally compartmentalized Viral components must be synthesized in different intracellular compartments

E N D
Lecture 18: Intracellular transport Flint et al., Chapter 12.
General remarks • Viruses are cellular parasites • Eukaryotic cells are functionally compartmentalized Viral components must be synthesized in different intracellular compartments • To assemble mature virions, virus components must be brought together for assembly from the different compartments. • Intracellular trafficking/sorting of viral nucleic acids, proteins, and glycoproteins is an essential prelude to virus particle assembly.
Cellular compartments: considerations • Nucleus • Cyotoplasm • Plasma membrane • Internal membranes • Mitochondria and other organelles • Viruses have evolved to assemble wherever a compartment exists
Virus assembly within the nucleus • Most DNA viruses assemble at the site of genome replication, the nucleus. • Viral structural proteins enter vial normal cellular pathway of nuclear protein transport
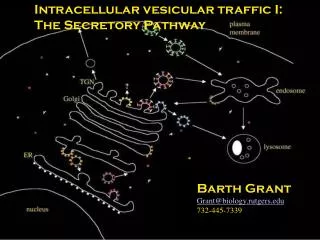
Envelopes of many viruses are derived from internal membranes Majority assemble at the cytoplasmic faces of compartments of the secretory pathway Bottom line: where there’s a membrane, there’s a place for enveloped viruses to assemble Interactions with internal cellular membranes Fig. 12.20
Virus assembly at the plasma membrane Enveloped viruses generally assembled at the plasma membrane. Integral membrane proteins are first transported to plasma membrane Internal proteins and nucleic acid genomes must also be actively transported to site of assembly (Fig. 12.2)
Intracellular trafficking: Considerations • Relative sizes of viruses and host cells: • Virus: diameters on the nm scale • Cells: diameters on the mm scale • Cell vol. 103 – 104 x greater than virus. • Viral components can’t find one another by passive diffusion. • Therefore, viral components must be actively transported to assembly sites.
Directional movement requires energy. Directional movement is generally achieved by: Protein channels (transporters, translocons, pores, portals) Move molecules across membranes Plasma Nuclear ER Golgi Mitochondria Motor proteins/cytoskeletal tracks Directional movement within the cytoplasm Myosin motors move cargo on actin fibers Dynein and kinesin motors move cargo on microtubules Getting from point A to point B. (Box 12.1)
Viral membrane proteins travel to the cell surface through a series of membrane-bound compartments and vesicles. ER to Golgi to plasma membrane. Trafficking requires cellular and viral proteins Vesicular transport to the cell surface (Fig. 12.9)
Transport of viral membrane proteins to the plasma membrane • They hitch rides on the cellular secretory pathway: • ER cis Goli network Golgi cisternae trans Golgi network plasma membrane • Maturation of proteins and structures occurs sequentially in different components. • Oligomerization occurs in ER and cis Golgi network • Oligosaccharides are trimmed, and further modified in Golgi cisternae • Proteins can be cleaved and further modified in trans Golgi network • Transported from trans Golgi to plasma membrane.
Snare proteins: ensuring targeting specificity • Snare proteins bring membranes close together • v-Snare: vesicular Snare • t-Snare: target Snare. • v- and t-Snares are highly specific for one another • Come together, bring vesicle and target membranes in close proximity to one another • Exclusion of water lowers energy barrier to membrane fusion • Upon fusion, contents of vesicle emptied into target area (Fig. 12.11)
Translocation of viral membrane proteins into the ER • Integral membrane proteins first inserted into ER membrane • Ribosomes targeted to ER by Signal Recognition Protein (SRP) • SRP interacts with ER bound SRP receptor • Directs ribosome to Translocation channel • Hydrophobic signal peptide associates with translocation channel • Remainder of protein translocated into ER lumen • Signal peptides cleaved off by signal peptidase • Hydrophobic Stop Signals used to stop translocation of protein into ER lumen (Box 12.2) • Results in transmembrane proteins
Translocation of viral membrane proteins into the ER (Fig. 12.6)
Hydrophobic Stop Signals used to stop translocation of protein into ER lumen Results in transmembrane proteins (Box 12.2)
Reactions within the ER lumen • Proteins can be modified within the ER lumen. • Glycosylation (modification with sugars) • Disulfide bond formation (between Cys residues) • Protein refolding by molecular chaperones • Oligomerization – most viral membrane proteins are complexed with one another
Reactions within the Golgi apparatus (Fig. 12.12)
Proteolytic cleavages occur in Golgi e.g. retroviral TM (transmembrane) and SU (surface unit) subunits of Env Fig. 12.13
Maturation of Influenza HA0 protein during transit through the secretory pathwayFig. 12.4
Viral genome transport • Viral genomes must also be transported to sites of virus particle assembly • Sequence specific viral proteins interact with viral genomes • These interact with the cellular transport systems to ensure delivery of genomes to the proper compartment
Viral inhibition/alteration of intracellular transport • It is often in the interest of the virus to inhibit components of the transport machinery • Many viruses inhibit transport of MHC class I molecules: prevents cell from signaling to the immune system that it is infected.
Some viruses inhibit transport beyond the compartment where they assemble. e.g. Poliovirus assembles in the ER. Poliovirus 2BC and 3A proteins inhibit vesicular transport beyond the ER. Force accumulation of virus components in the compartment where they assemble. Viral inhibition/alteration of intracellular transport Fig. 12.16
Viral inhibition/alteration of intracellular transport • Some viruses redirect transport system to decrease expression of their target proteins on the plasma membrane. • e.g. HIV Nef and Vpu redirect CD4 away from plasma membrane. • This helps to ensure that new virus won’t fuse back into infected cell.