Gene Regulation and Development
790 likes | 1.05k Vues
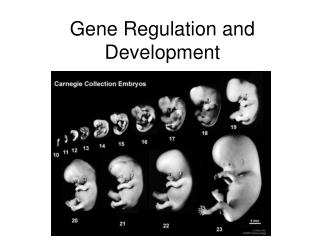
Gene Regulation and Development. Making of the Complex Living things. In the development of multicell organisms, a single-cell zygote gives rise to many different cell types.

Gene Regulation and Development
E N D
Presentation Transcript
Making of the Complex Living things • In the development of multicell organisms, a single-cell zygote gives rise to many different cell types. • Each type has different structure, under taking different chemical and biologic reactions and performs corresponding functions. • All these events are controlled by sets of genes in the cell and influenced by surrounding stimuli. • Body system of multicell organisms • Cells of similar types are organized into tissues • Tissues with associated function into organs • Organs of associated function into organ systems • Organ systems coordinately to support the whole organism • Process of embryonic development must give rise not only to the cells of different types but also to higher-level structures and coordinated functions.
How life begin… • An organism arises from a fertilized egg as the result of three interrelated processes: cell division, cell differentiation, and cell morphogenesis. • During development, cells becomes polarized, starts divide unevenly, reprograms gene expression and become specialized in structure and function, a process called differentiation. • The processes of morphogenesis is forming of shapes, give cells, tissues and organs their proper shape to fit their corresponding functions. • All these processes are part of development.
Embryonic Development Fertilization results in a zygote and triggers embryonic development Middlepiece Neck Head Plasma membrane Tail Mitochondrion(spiral shape) Nucleus Acrosome
Only one sperm willpenetrate this human egg to initiate the fertilization There is a selection of sperm quality 1 The spermapproachesthe egg 2 The sperm’sacrosomal enzymesdigest the egg’s jelly coat 3 Proteins on thesperm head bindto egg receptors SPERM 4 The plasma membranesof sperm and egg fuse Spermhead The spermnucleus enters the egg cytoplasm 5 Nucleus Acrosome Acrosomalenzymes Plasmamembrane 6 Afertilizationenvelopeforms Receptor protein molecules Plasmamembrane Spermnucleus Vitellinelayer Cytoplasm Eggnucleus Jellycoat (Zona Pellucida) The nucleiof spermand egg fuse 7 EGG CELL Zygotenucleus
In mammals, fertilization occurs in the oviduct.Sperm fuses with oocyte in metafase2, completes meiosis and then two parenteral pronuclei fuse to form the diploid zygote. Cleavage starts Fertilizationof ovum Ovary Oviduct Secondaryoocyte Blastocyst(implanted) Endometrium Ovulation Uterus
Cell Division Cleavage is the first major step of embryonic development It is the rapid succession of cell divisions It creates a multicellular embryo It partitions the multicellular embryo into developmental regions Cells and embryo become polarized Cleavageis Controlled by cyclins and cyclin-dependent kinases (Cdks) Duringcleavage,the zygote is divided into smaller & smaller cells called blastomeres Embryo volume does not change
Non-differentiated cells and can give rise to any tissue. Stem cells continue to divide and remain undifferentiated. Totipotent: cells that can give rise to any cell types Pluripotent: cells that can give rise to multiple cell types Tissue-specific: can give rise to only one or fewer cell types Blastomeres
Cell divide and cell number increase continueously for 5-6 days(3-4 days in mice) to produce the first morphologically differentiated structure The first two different cell types with totally different gene expression patterns The blastocyst consists of: Outer layer(Trophoblast cell )_which secretes enzymes to enable the blastocyst to implant on endometrium surface and laterforms placenta Inner layer (Inner Cell Mass, which is undifferentiated and a general source of embryonic stem cell)_which develop to complete body parts. Blastocyst ENDOMETRIUM Inner cell mass Trophoblast Cavity Antibody against Oct4 Trophoblast
ES Cell Culture Once sperm and egg cell fuse (zygote), cell start to divide anddifferentiate. The first differentiated structure: a blastocyst. The inner cell massof the blastocyst develops into embryo. Embryonic stem-cell culture Inner cell mass Egg Embryonic stem cells (ES cells) are isolated from the inner cell mass Sperm Blastocyst Embryo
Determination • The earliest changes that set a cell to a specialized stage without turning back at molecular level. • Can only be “seen” by experiment • Cells occupy a specific location in the embryo • They are not “naturally” totipotent any more • Molecular changes that drives the process in the embryo is termed determination, the beginning of differentiation. • During embryonic development, cells become obviously different in structure and function as they differentiate.
Cell Differentiation Gene expression is switched to a new program Cells initiate developmental changes by using new transcriptional factors and give a totally different pattern of gene expression Cells become committed to a particular developmental pathway: via genetic program under the differential inheritance ofcytoplasmic determinants via environmental cell-cell interactions Cells are “morphologically” distinguishable by different size and shape
Differentiated Cells • These cells produce the proteins that allow them to carry out their specialized roles in the organism. • For example, only lens cells devote 80% of their capacity for protein synthesis to making just one type of proteins, crystallins. • These form transparent fibers that allow the lens to transmit and focus light. • Similarly, skeletal muscles cells have high concentrations of proteins specific to muscle tissues, such as muscle-specific version of the contractile protein myosin and the structural protein actin. • They also have membrane receptor proteins that detect signals from nerve cells.
Morphogenesis Polar Cell division The orientation of the mitotic spindle determines the plane of cell division in eukaryotic cells -If spindle is centrally located, two equal-sized daughter cells will result -If spindle is off to one side, two unequal daughter cells will result Cell shape and size In animals, cell differentiation is accomplished by profound changes in cell size and shape -Nerve cells develop long processes called axons -Skeletal muscles cells are large and multinucleated Cell death -Necrosis is accidental cell death in group, not cell-specific -Apoptosisis programmed cell death, can happen in individual cells -Is required for normal development in all animals -“Death program” pathway consists of: -Activator, inhibitor and apoptotic protease Cell migration -Cell movement involves adhesion and loss of adhesion between cells and substrate -Cell-to-cell interactions are often mediated through cadherins -Cell-to-substrate interactions often involve complexes between integrinsand theextracellular matrix (ECM)
Gastrulation produces a three-layered embryo The second major step of embryonic development, produces a three-layered embryo It happens after implantation The size of embryos starts to grow It adds more cells and volume to the embryo It sorts all cells into three distinct cell layers The embryo is transformed from the blastula into the gastrula The three layers Ectoderm, the outer layer Endoderm, an inner layer, give rise to the embryonic digestive tract Mesoderm, which partly fills the space between the ectoderm and endoderm
Embryonic tissue layers begin to develop into specific tissues and organ systems after gastrulation
The embryo floats in amniotic cavity, chorion and embryonic mesodermforms the placenta • Functions of thechorion: • Provide nutrients and O2to the fetus • Secrete hormones into themother to help retain thefetus • Repress the mother’simmune response to preventrejection of the fetus • The placenta’s chorionic villi absorb food and oxygen from the mother’s blood Placenta Mother’s bloodvessels Amnioticcavity Allantois Yolk sac Embryo Chorion Chorionicvilli
Embryonic induction initiates organ formation Induction is the mechanism by which one group of cells influences the development of tissues and organs from ectoderm, endoderm, and mesoderm Adjacent cells and cell layers use chemical signals to influence differentiation Chemical signals turn on a set of genes in the receiving cells and leads to differentiation into a specific tissues from these cells Pattern formation Pattern formation is the emergence of structures in their correct relative positions It often involves the response of cells and their genes to spatial gradient of chemicals
Induction: Tissues and organs take shape in a developing embryo as a result of Cell division Cell shape changes Cell migration Programmed cell death (apoptosis) Ectoderm
Pattern Formation Differentiation is carefully orchestrated: • Proliferation • Cell migration • Interactions (Induction) • Epithelial-mesenchymal transformations • Epithelial folding, movement, evagination, fusion • Apoptosis …
Apoptosis • Lineage analysis of C. elegans highlights another outcome of cell signaling, programmed cell death or apoptosis. • The timely suicide of cells occurs exactly 131 times in the course of C. elegans’s normal development. • At precisely the same points in development, signals trigger the activation of a cascade of “suicide” proteins in the cells destined to die.
Organism Caenorhabditis elegans Mammalian Cell Inhibitor: CED-9 Inhibitor Bcl-2 Activator: CED-4 Apaf1 Apoptotic Protease: CED-3 Caspase-8 or -9 Inhibition Apoptosis Apoptosis Activation a. b.
If it weren’t for apoptosis, you may look a little different… • Apoptosis pathways in humans and other mammals are more complicated. • Research on mammals have revealed a prominent role for mitochondria in apoptosis. • Signals from apoptosis pathways or others somehow cause the outer mitochondrial membrane to leak, releasing proteins that promote apoptosis. • A cell must make a life-or-death “decision” by somehow integrating both the “death” and “life” (growth factor) signals that it receives. • Apoptosis is essential to the development of animal morphogenesis (prevents webbing between fingers and toes).
Model Organism (1)The Worm • The nematode C. elegans normally lives in the soil but is easily grown in petri dishes. • Only a millimeter long, it has a simple, transparent body with only a few cell types and grows from zygote to mature adult in three and a half days. • Its genome has been sequenced. • Because individuals are hermaphrodites, it is easy to detect recessive mutations. • Self-fertilization of heterozygotes will produce some homozygous recessive offspring with mutant phenotypes. • A further important feature is that every adult C. elegans have exactly 959 somatic cells. • These arise from the zygote in virtually the same way for every individual.
Studying Worms • C. elegans is a very useful model organism for investigating the roles of cell signaling and induction in development. • The pathway from fertilized egg to adult nematode involves four larval stages (the larvae look much like smaller versions of the adult) during which the vulva develops. • Six cells present on the ventral surface of the second-stage larva gives the vulva. • A single cell in the embryonic gonad, the anchor cell, initiates a cascade of signals that establishes the fate of the vulva precursor cells. • Vulva development in the nematode illustrates several important developmental concepts: • In the developing embryo, sequential inductions drive the formation of organs. • The effect of an inducer can depend on its concentration. • Inducers produce their effects via signal-transduction pathways similar to those operating in adult cells. • The induced cell’s response is often the activation (or inactivation) of genes which establishes the pattern of gene activity characteristic of a particular cell type. • Genetics as a powerful approach for elucidating the mechanisms of development.
Model Organism (2)The Fruit Fly • The fruit fly Drosophila melanogaster was first chosen as a model organism by geneticist T.H. Morgan and intensively studied by generations of geneticists. • The fruit fly is small and easily grown in the laboratory. • It has a generation time of only two weeks and produces many offspring. • Embryos develop outside the mother’s body. • In addition, there are vast amounts of information on its genes and other aspects of its biology. • However, because first rounds of mitosis occurs without cytokinesis, parts of its development are superficially quite different from what is seen in other organisms.
Sabotaging the Fruit Flies • In the 1940s, Edward B. Lewis demonstrated that the study of mutants could be used to investigate Drosophila development. • He studied bizarre developmental mutations and located the mutations on the fly’s genetic map. • This research provided the first concrete evidence that genes somehow direct the developmental process. • In the late 1970s, Christiane Nüsslein-Volhard and Eric Weischaus pushed the understanding of early pattern formation to the molecular level. • Their goal was to identify all the genes that affect segmentation in Drosophila. • Mutations that affect segmentation are likely to be embryonic lethals, leading to death at the embryonic or larval stage. • Because of maternal effects on axis formation in the egg, they also needed to study maternal genes. • After a year of hard work, they identified 1,200 (out of a fruit flies 13,000) genes essential for embryonic development • About 120 of these were essential for pattern formation leading to normal segmentation. They were able to map these genes. • The results of detailed anatomical observations of development in several species and experimental manipulations of embryonic tissues laid the groundwork for understanding the mechanisms of development.
Homeotic-Selector HOX Genes
Background Among the most fascinating kinds of abnormalities in animals are those in which one normal body part is replaced by another. William Bateson (1894) catalogued several oddities of this nature coining the term homeotic to describe them. Calvin Bridges (1915) isolated a spontaneous mutant of Drosophila in which part of the haltere was transformed into wing tissue. The mutant was called bithorax (duplication of a thoracic segment). In the following decades other mutations affecting segment identity were identified e.g. certain Antennapedia mutations causing transformation of antennae into legs. In Drosophila, genes controlling segment identity were found to be transcription factors and coined to be homeotic-selector genes, meaning that the fate of a given segment was selected by the expression of specific homeotic genes.
Homeobox • All homeotic genes of Drosophila include a 180-nucleotide sequence called the homeobox, which specifies a 60-amino-acid homeodomain. • An identical or very similar sequence of nucleotides (often called Hox genes) are found in many other animals, including humans.
How Homeodomains Work • Proteins with homeodomains probably regulate development by coordinating the transcription of batteries of developmental genes. • In Drosophila, different combinations of homeobox genes are active in different parts of the embryo and at different times, leading to pattern formation.
Action of Signaling molecules Paracrine factors • Inducing factors controlling forms of developing organ • Mitogen regulating cell proliferation • Morphogen acting in a dose-dependent way to pattern the cell fates within a target field
Morphogens • Product of the mother’s bicoid gene is essential for setting up the anterior end of the fly. • The gene’s products are concentrated at the future anterior end. • This is a specific version of a general gradient hypothesis, in which gradients of morphogens establish an embryo’s axes and other features. • The bicoid protein and other morphogens are transcription factors that regulate the activity of some of the embryo’s own genes. • Gradients of these morphogens bring about regional differences in the expression of segmentation genes, the genes that direct the actual formation of segments after the embryo’s major axes are defined.
Homeotic Mutations The direction of homeotic transformations depends on whether the mutation causes loss of homeotic gene function where the gene normally acts or gain of function where the gene normally does not act. Ultrabithorax (Ubx) acts in the haltere to promote haltere development and repress wing development. Loss of function mutations in Ubx transform the haltere into a wing. Dominant mutations that cause Ubx to gain function in the wing transform that structure into a haltere. In antenna-to-leg transformations of Antennapedia the mutants reflect a dominant gain of Antennapedia gene function in the antennae.
Normal adult fly Antennapedia mutant Bithorax mutant Examples of Homeotic Mutations
Homeotic-Selector Genes • Homeotic gene products regulate development within parasegmental domains. At this stage each band of cells along the A-P axis express a unique combination of transcription factors which control subsequent cell development. • Patterns of selector gene expression are determined in the early embryo and maintained into the adult fly. Continuous expression is required to specify structures along the A-P axis. • The genes are ordered on the chromosome in the order of their expression with respect to the parasegments of the Drosophila embryo. • Two of the best studied selector genes are Antennapaedia(Antp) which dominates development of the fourth parasegment and Ultrabithorax (Ubx) which largely controls the sixth parasegment.
Homeotic-Selector Genes • Transcription of the Antp and Ubx genes begins in the third hour of embryogenesis in domains determined in part by various gap-gene products with boundaries set up by the segmentation genes. • Homeotic genes act in a cell autonomous manner – a cell will behave according to homeotic gene expression of its own regardless of the pattern of expression of its neighbours • Maintenance of homeotic gene expression is through self regulation and the Polycomb-like class of genes. • Selector genes specify epidermal structures – the musculature, neural tissue and gut along the A-P axis. • Two homeotic selector gene clusters occur in two groups on the same chromosome – five (the Antennapedia Complex – ANT-C) are expressed in the head region while genes in the Bithorax Complex (BX-C) are mainly expressed in thorax or abdominal regions.
Eight Genes Regulate the Identity of Within the Adult and Embryo labial (lab) proboscipedia (pb) Deformed (Dfd) Sex combs reduced (Scr) Antennapedia (Antp) Ultrabithorax (Ubx) abdominal A (abd-A) Abdominal B (Abd-B)
Homeotic-Selector Genes • Homeotic genes encode nuclear proteins containing a DNA-binding motif called a homeodomain. • The products are transcription factors that specify segment identity by activating multiple gene expression events. • The genes are initially activated imprecisely by the concentration gradients of gap gene products. e.g. Ubx is switched on between certain concentrations of hunchback to give a broad band of expression near the middle of the embryo. Later, fushi tarazu and even skipped sharpen the limits of Ubx expression which comes into register with the anterior boundaries of specific parasegments. • The BX-C and ANT-C genes have extensive non-coding sequences (introns) that are critical in regulating their individual expression.
Fate map of a Drosophila embryo at the cellular blastoderm stage. • The embryo is shown in side view and in cross-section, displaying the relationship between the dorsoventral subdivision into future major tissue types and the anteroposterior pattern of future segments. • A heavy line encloses the region that will form segmental structures. • During gastrulation the cells along the ventral midline invaginate to form mesoderm, while the cells fated to form the gut invaginate near each end of the embryo. • Thus, with respect to their role in gut formation, the opposite ends of the embryo, although far apart in space, are close in function and in final fate.
Patterns of Expression • The patterns of expression compared to the chromosomal locations of the genes of the HOM complex. • The sequence of genes in each of the two subdivisions of the chromosomal complex corresponds to the spatial sequence in which the genes are expressed. • Note that most of the genes are expressed at a high level throughout one parasegment (dark color) and at a lower level in some adjacent parasegments (medium color) where the presence of the transcripts is necessary for a normal phenotype, light color where it is not). • In regions where the expression domains overlap, it is usually the most "posterior" of the locally active genes that determines the local phenotype. • The drawings in the lower part of the figure represent the gene expression patterns in embryos at the extended germ band stage, about 5 hours after fertilization.
Effects of Mutations in Bithorax Complex • Contribution of BX-C genes — Ubx, abdA, and AbdB—to determination of parasegment identity. • The numbers above each larva indicate the parasegments; those below, the corresponding segments. • The cuticular pattern of larvae is used to assign an identity to each parasegment (PS), which is indicated by color, as depicted in the wild type at the top. • Red PS and segment labels indicate abnormal patterns that do not correspond exactly to any found in wild-type larvae.
Expression and lethal embryonic phenotypes of homeotic genes. • This phenotypic analysis has been confirmed by the expression patterns of the various genes in various mutant backgrounds. • The anterior limits of the domain of expression of a particular homeotic gene are presumably set by a collaboration between gap genes and pair-rule genes. • Removal of an anteriorly-acting homeotic has no effect on expression or phenotype in the domain of a more posteriorly acting homeotic gene. • Antp expands posteriorly from PS4 to PS6 in a Ubx mutant • Antp expands posteriorly from PS4 to PS12 in a Ubx AbdA mutant • Antp expands posteriorly from PS4 to PS14 in a Ubx AbdA AbdB mutant • Ubx expands posteriorly from PS6 to PS12 in a AbdA mutant • Ubx expands posteriorly from PS6 to PS14 in a AbdA AbdB mutant
Gain- and Loss-of-Function Studies • Conservation of order of homeotic gene expression plays an important role in controlling patterning. • “Out of order” expression in single mutants lacking abd-A causes marked defects in parasegments 10-14 and they do not correspond to any wild-type parasegment. Therefore Ubx and Abd-B do not provide recognisable patterning information in the absence of abd-A. • For gain of function, transfected Ubx can be induced to be uniformly expressed along the A-P axis (as opposed to parasegments 5 and 6). In this scenario parasegments 6 to 14 form normally but parasegments 1 to 5 are transformed into parasegment 6. Gain- and loss-of-function studies show a consistent relation between selector genes: Genes expressed more posteriorly suppress the action of genes expressed more anteriorly.
Binding Sites of Hox Proteins • Hox proteins bind DNA sequences – but different Hox proteins bind with high affinity to the same short DNA sequences found once every kilobase. • The ability of Hox proteins to control expression of different genes depends on the product of the extradenticle (exd) gene. • Exd binds different Hox proteins forming different heterodimers that bind selectively to specific target sequences • Exd may also contribute to specificity of Hox function by converting bound Hox proteins from repressors to activators. • Exd may also operate independently to repress other genes. • Role of Exd protein in conferring DNA-binding specificity on Drosophila Hox proteins. • Various Hox proteins, including Dfd and Lab, bind to a 10-bp consensus sequence that differs in the nucleotides (N) at the two central positions. • (b) Exd-Dfd and Exd-Lab heterodimers specifically recognize Hox-binding sites in which the central dinucleotide is TA or GG, respectively.
Maintaining Hox Gene Expression • The transcription-control regions of some Hox genes contain binding sites for their encoded proteins – autoregulatory loop (e.g. lab and Dfd). • A second mechanism requires proteins that modulate chromatin structure. There are two classes – the trithorax groupand the polycomb group. • Early patterning requires repression as well as activation of gene expression. Polycomb proteins have a repressive effect on the expression of Hox genes. • Polycomb proteins bind multiple chromosomal locations to form large macromolecular complexes and this becomes “locked in”. • Trithorax proteins maintain the expression of many Hox genes. These also form large multiprotein complexes at multiple chromosomal sites, but mainain an open chromatin structure and stimulate gene expression.
Pattern of Binding of Polycomb Protein The normal pattern of binding of Polycomb protein to Drosophila giant chromosomes, visualized with an antibody against Polycomb. The protein is bound to the Antennapedia complex (ANT-C) and the bithorax complex (BX-C) as well as about 60 other sites.
Conservation of Homeotic-Selector Genes • The organisation of genes within ANT-C and BX-C are similar to mammalian homologues occur in four gene clusters located on different chromosomes (Hox complex). • Current thinking on the evolution is as follows: • A primordial homeotic selector gene of a common ancestor of worms, flies and vertebrates underwent repeated duplication to form a series of genes in tandem. • In the Drosophila sublineage this single complex became split into separate Antennapedia and bithorax complexes. • In the lineage leading to the mammals the whole complex was repeatedly duplicated to give the four Hox complexes. • The parallelism between mammals and Drosophila is not perfect as some individual genes have been duplicated and others lost since the complexes diverged.