Translation
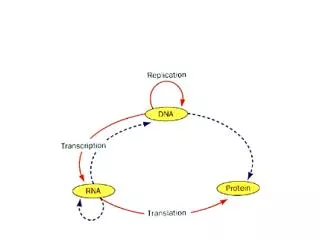
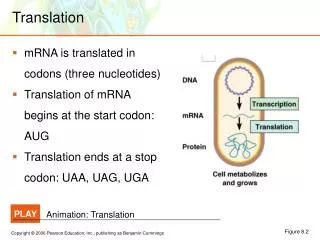
Translation. Questions? 1) How does poliovirus shutoff eukaryotic translation? If eukaryotic messages are not translated how can poliovirus get its message translated? Host Cell Shutoff Initiation of eukaryotic translation involves many initiation factors


Translation
E N D
Presentation Transcript
Translation • Questions? 1) How does poliovirus shutoff eukaryotic translation? • If eukaryotic messages are not translated how can poliovirus get its message translated? Host Cell Shutoff • Initiation of eukaryotic translation involves many initiation factors 2) Pathway for initiation is more complicated than for prokaryotic translation a) initiator met tRNA binds to the 40S subunit in the presence of eIF-2 b) eIF-4F + GTP enter the complex and recognize a capped mRNA c) the complex then ‘scans’ down the RNA until an AUG is found d) eIF-5 binds as eIF-2 leaves allowing the 60S subunit to bind and the peptide bond formation starts
Translation (cont’d) 3) eIF-4F is involved in cap recognition and mRNA binding (270000D) a) made up of three subunits i) cap binding protein (also known as eIF-4E, 25000D) ii) ATP-dependent RNA helicase, removes secondary structure from the 5' end of mRNAs (also known as eIF-4A, 25000D) iii) p220, or eIF-4G, function unknown binds to eIF-4A and -4E b) p220 is cleaved in poliovirus-infected cells i) direct analysis of cleaved products ii) purified eIF4-F can restore cap-dependent translation iii) poliovirus-infected cells can contribute all components to a fractionated in vitrotranslation system except eIF-4F c) cap-independent translation can occur d) viral-proteinase responsible? i) antibodies to 3C to not inhibit cleavage ii) 3C cleavage activity does not co-purify with p220 cleavage iii) translation of 2A by itself leads to p220 cleavage
Translation (cont’d) • Ok, so how does polio get its RNA translated? 1) polio has a 5' nontranslated region of ~750 nt before the AUG that initiates the polyprotein; this region contains 7 unused AUGs 2) mutations in the 5' nontranslated region can have a dramatic affect on pathogenesis 3) internal ribosome binding has been shown to occur using bicistronic mRNA 4) the internal entry is insensitive to cap analog inhibition (cap-independent) 5) this sequence has been called: IRES (internal ribosome binding site) 6) these sequences are ~400 to 500 nts in length and consist of a series of highly ordered secondary structures
Cellular Proteins? • What are the cellular proteins that bind to this sequence? 1) Currently there are no known eIF that bind 2) there is a highly conserved polypyrimidine tract that lies either ~20nt before the initiator AUG (EMC) or ~175 nt before the initiator AUG (polio) 3) p52 binds to this tract as determined by crosslinking studies i) p52 is the La autoantigen (patients with lupus have ab) ii) La is known to bind 3' oligouridylates on RNA pol III transcripts iii) it is believed to play a role in RNA pol III transcription termination iV) purified La stimulates poliovirus translation in vitro 4) p57 binds to near stem loop V and VI i) p57 is the pyrimidine-tract binding protein (PTB) ii) has been implicated in nuclear processes involving pre-mRNA iii) immunodepletion of PTB inhibits polio and EMC translation but not globin 5) poly (rC) binding protein 5) What are nuclear proteins doing in the cytoplasm and why are they helping viruses? 6) There are several examples of internal initiation for several cellular messages: a) antennapedia mRNA b) fibroblast growth factor mRNA c) Bip mRNA
Assembly 1) Proteolytic processing of the P1 proteins a. cleavage of the P1/2A cleavage site by 2Apro b. cleavage of P1 by 3CD: generates three viral coat proteins VP1, VP3, VP0 (VP4 + VP2; this cleavage occurs later in assembly) 2) These three viral proteins form the basic building block of the virus termed the protomer (smallest identical subunit of an oligomeric protein). The protomer exists in infected cells and is described by its sedimentaion value: the 5S particle 3) 5 protomers assemble to form a pentamer or 14S particle 4) 12 pentamers assemble to form either: a. procapsid (a 70S empty capsid, ie no RNA) that is subsequently packaged with viral RNA to form a provirion b. provirion - a complete particle that must undergo a maturation cleavage to generate infectious 155S virions 5) Maturation cleavage: a. cleavage of VP0 into VP2 + VP4 b. mechanism unclear, RNA probably required
Atomic Structure of the Protomer 1) protomer is wedge-shaped with VP1, VP2, VP3 occupying separate domains on the outer surface, VP4 is internal 2) VP1, VP2, VP3 structurally similar with one another 3) Fold consists of 8 antiparallel beta barrels - has been seen with every RNA virus capsid protein seen so far (except Sindbis) 4) Loops connecting beta strands form some of the protruding surface features seen in the virus, most of the neutralizing immunogenic sites (NIM) are found in these loops 5) protomers are connected at the pentamer vertex through the amino-terminal tails of VP3 -> they form a twisted tube called the beta annulus 6) protomers are connected together by intertwining of VP1, VP3, and VP0 N and C tails, once VP0 undergoes cleavage VP2 is only loosely associated
Receptors Receptors: Function as attachment sites for a virus, the receptor may or may not be involved in virus / nucleic acid penetration. There are many ways to isolate a cellular receptor for a virus, usually one starts with: 1) an antibody directed against the cellular protein How do you get this? a. injection of cells into mice to generate monoclonals that recognize the receptor b. purify protein by affinity to virus and generate antibodies 2) a receptor negative cell line (virus insensitive cell line) a. transfection of cDNA from a receptor positive cell line can transform such cells from insensitive to virus sensitive cells b. transfection of cDNA from a receptor positive cell line can be screened with a monoclonal antibody against the receptor protein
Entry I 1) Binding of virus to the cell surface causes a conformational change in the virion resulting in the release of VP4 and the conversion of the infectious particle to a non-infectious A-particle 2) Binding of poliovirus to soluble receptor results in conversion of infectious particle to the non-infectious A-particle 3) Observations (1) and (2) suggest a pathway for entry: following binding of virus particles to the receptor, conformational changes occur which result in the release of the internal VP4 molecules and positioning of the hydrophobic N-termini of VP1 into the membrane forming a pore by which the genome RNA passes through.