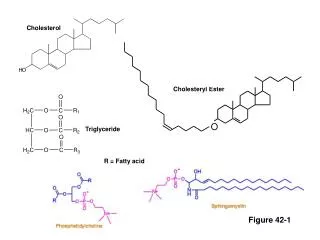
D- Cholesterol Synthesis
D- Cholesterol Synthesis. Hydroxymethylglutaryl-coenzyme A ( HMG-CoA ) is the precursor for cholesterol synthesis. HMG-CoA is also an intermediate on the pathway for synthesis of ketone bodies from acetyl-CoA.


D- Cholesterol Synthesis
E N D
Presentation Transcript
Hydroxymethylglutaryl-coenzyme A (HMG-CoA) is the precursor for cholesterol synthesis. HMG-CoA is also an intermediate on the pathway for synthesis of ketone bodies from acetyl-CoA. The enzymes for ketone body production are located in the mitochondrial matrix. HMG-CoA destined for cholesterol synthesis is made by equivalent, but different, enzymes in the cytosol.
HMG-CoAis formed by condensation of acetyl-CoA & acetoacetyl-CoA, catalyzed by HMG-CoA Synthase. • HMG-CoA Reductase catalyzes production of mevalonate from HMG-CoA.
The carboxyl of HMG that is in ester linkage to the CoA thiol is reduced to an aldehyde, and then to an alcohol. NADPH serves as reductant in the 2-step reaction. Mevaldehyde is thought to be an active site intermediate, following the first reduction and release of CoA.
HMG-CoA Reductase is an integral protein of endoplasmic reticulum membranes. The catalytic domain of this enzyme remains active following cleavage from the transmembrane portion of the enzyme. The HMG-CoA Reductase reaction, in which mevalonate is formed from HMG-CoA, is rate-limiting for cholesterol synthesis. This enzyme is highly regulated and the target of pharmaceutical intervention.
Mevalonate is phosphorylated by 2 sequential Pi transfers from ATP, yielding the pyrophosphate derivative. ATP-dependent decarboxylation, with dehydration, yields isopentenyl pyrophosphate.
Isopentenyl pyrophosphate is the first of several compounds in the pathway that are referred to as isoprenoids, by reference to the compound isoprene.
Isopentenyl Pyrophosphate Isomeraseinter-converts isopentenyl pyrophosphate & dimethylallyl pyrophosphate. Mechanism: protonation followed by deprotonation.
Condensation Reactions Prenyl Transferase catalyzes head-to-tail condensations: • Dimethylallyl pyrophosphate & isopentenyl pyrophosphate react to form geranyl pyrophosphate. • Condensation with another isopentenyl pyrophosphate yields farnesyl pyrophosphate. • Each condensation reaction is thought to involve a reactive carbocation formed as PPiis eliminated.
Each condensation involves a carbocation formed as PPi is eliminated.
Squalene Synthase: Head-to-head condensation of 2 farnesyl pyrophosphate, with reduction by NADPH, yields squalene.
Squaline epoxidase catalyzes conversion of squalene to 2,3-oxidosqualene. This mixed function oxidation requires NADPH as reductant & O2 as oxidant. One O atom is incorporated into substrate (as the epoxide) & the other O is reduced to water.
Squalene Oxidocyclase catalyzes a series of electron shifts, initiated by protonation of the epoxide, resulting in cyclization. Structural studies of a related bacterial enzyme have confirmed that the substrate binds at the active site in a conformation that permits cyclization with only modest changes in position as the reaction proceeds. The product is the sterol lanosterol.
Conversion of lanosterol to cholesterol involves 19 reactions, catalyzed by enzymes in ER membranes. Additional modifications yield the various steroid hormones or vitamin D.
Many of the reactions involved in converting lanosterol to cholesterol and other steroids are catalyzed by members of the cytochrome P450enzyme superfamily. The human genome encodes 57 members of the cyt P450 superfamily, with tissue-specific expression and intracellular localization highly regulated. • Some P450 enzymes are localized in mitochondria. • Others are associated with endoplasmic reticulum membranes.
Cyt P450 enzymes catalyze various oxidative reactions. Many are mixed function oxidations(mono-oxygenations) that require O2& a reductant,e.g.,NADPH. One oxygen atom is incorporated into a substrate & the other oxygen atom is reduced to water. An example is hydroxylation of a steroid as in the ER electron transfer pathway above: NADPH transfers 2 electrons to cytochrome P450via a reductase that has FAD & FMN prosthetic groups.
A cysteine S atom typically serves as an axial ligand (X or Y) for the iron atom of a cyt P450heme. The other axial position, where O2 binds, may be open or have a bound H2O that is displaced by O2.
O2 is cleaved after binding to the reduced P450 heme iron. • In the example shown: • one oxygen atom is reduced to water • and a substrate is hydroxylated.
Reactions catalyzed by different P450 enzymes include hydroxylation, epoxidation, dealkylation, peroxidation, deamination, desulfuration, dehalogenation, etc. • P450 substrates include steroids, polyunsaturated fatty acids, eicosanoids, retinoids, & various non-polar xenobiotics (drugs & other foreign compounds). Some P450 enzymes have broad substrate specificity. • Mechanisms for detoxification of non-polar compounds include reactions such as hydroxylations that increase polarity, so that the products of these reactions can be excreted by the kidneys. Explore with Chime the hemoprotein domain of a Bacillus magaterium cytochrome P450.
Farnesyl pyrophosphate, an intermediate on the pathway for cholesterol synthesis, also serves also as precursor for synthesis of various non-steroidal isoprenoids. The importance of the other products of the pathway that originates with mevalonate is reflected in serious diseases that result from genetic defects in this pathway.
Prenylated proteins have covalently linked geranylgeranyl or farnesyl groups that anchor them to membranes. Many proteins involved in cell signaling have such lipid anchors, including small GTP-binding proteins such as Ras.
Farnesyl Transferase catalyzes transfer of the farnesyl moiety of farnesyl pyrophosphate to a cysteine residue in a sequence CaaX at the C-terminus of a protein, "a" being an aliphatic amino acid. After subsequent cleavage of the terminal 3 amino acids, the new terminal carboxyl may be methylated, further increasing hydrophobicity.
Some other isoprenoids: • Dolichol pyrophosphate has a role in synthesis of oligosaccharide chains of glycoproteins. Additional roles have been proposed; dolichol is found in many membranes of cells.
Coenzyme Q (ubiquinone), which has an isoprenoid side-chain, functions in the electron transfer chain.
Heme a, a constituent of respiratory chain complexes, has a farnesyl side-chain.
Regulation of cholesterol synthesis HMG-CoA Reductase, the rate-limiting step on the pathway for synthesis of cholesterol, is a major control point. Regulation relating to cellular uptake of cholesterol will be discussed in the next class. Short-term regulation: HMG-CoA Reductase is inhibited by phosphorylation,catalyzed by AMP-Dependent Protein Kinase (which also regulates fatty acid synthesis and catabolism). This kinase is active when cellular AMP is high, corresponding to when ATP is low. Thus, when cellular ATP is low, energy is not expended in synthesizing cholesterol.
Long-term regulationis by varied formation and degradation of HMG-CoA Reductase and other enzymes of the pathway for synthesis of cholesterol. • Regulatedproteolysis of HMG-CoA Reductase: • Degradation of HMG-CoA Reductaseis stimulated by cholesterol, oxidized derivatives of cholesterol, mevalonate, & farnesol (dephosphorylated farnesyl pyrophosphate). • HMG-CoA Reductase includes a transmembrane sterol-sensing domain that has a role in activating degradation of the enzyme via the proteasome (proteasome to be discussed later).
Regulated transcription: • A family of transcription factors designated SREBP (sterol regulatory element binding proteins) regulate synthesis of cholesterol and fatty acids. Of these, SREBP-2 mainly regulates cholesterol synthesis. (SREBP-1c mainly regulates fatty acid synthesis.) • When sterol levels are low, SREBP-2 is released by cleavage of a membrane-bound precursor protein. • SREBP-2 activatestranscription of genes forHMG-CoA Reductase and other enzymes of the pathway for cholesterol synthesis.
The SREBP precursor protein is embedded in the endoplasmic reticulum (ER) membrane via two transmembrane α-helices. The N-terminal SREBP domain, which extends into the cytosol, has transcription factor capability. The C-terminal domain, also on the cytosolic side of the membrane, interacts with a cytosolic domain of another ER membrane protein SCAP (SREBP cleavage-activating protein).
SCAP has a transmembranesterol-sensing domainhomologous to that of HMG-CoA Reductase. When bound to a sterol, the sterol-sensing domain of SCAP binds the ER membrane protein Insig. Association with Insig causes the SREBP-SCAP precursor complex to be retained within the ER. When sterol levels are low, SCAP & Insig do not interact. This allows the SCAP-SREBP precursor complex to translocate from the ER to the golgi apparatus.
ProteaseS1P (site one protease), an integral protein of golgi membranes, cleaves the SREBP precursor at a site in the lumenal domain. • An intramembrane zinc metalloprotease domain of another golgi proteaseS2P then catalyzes cleavage within the transmembrane segment of the SREBP precursor, releasingSREBP to the cytosol. • Only the product of S1P cleavage can serve as a substrate for S2P.
The released SREBP enters the cell nucleus where it functions as a transcription factor to activate genes for enzymes of the cholesterol synthesis pathway. Its lifetime in the nucleus is brief, because SREBP is ubiquitinated & degraded. Diagram(in article by P. J. Espenshade; requires J. Cell Sci. subscription) Homodimeric DNA-binding domain of SREBP interacting with a sterol regulatory element DNA segment.
Drugs used to inhibit cholesterol synthesis include competitive inhibitors of HMG-CoA Reductase. Examples include various statin drugs such as lovastatin (Mevacor) and derivatives (e.g., Zocor), Lipitor, etc. A portion of each statin is analogous in structure to mevalonate or to the postulated mevaldehyde intermediate. Extensive clinical trials have shown that the statin drugs decrease blood cholesterol and diminish risk of cardiovascular disease.
Since farnesyl & geranylgeranyl membrane anchors are important for signal proteins that regulate cell cycle progression, inhibitors of prenylating enzymes such as Farnesyl Transferase are being tested as anti-cancer drugs. However, toxic side effects may limit usefulness of this approach.