Download
1 / 1
10 likes | 121 Vues
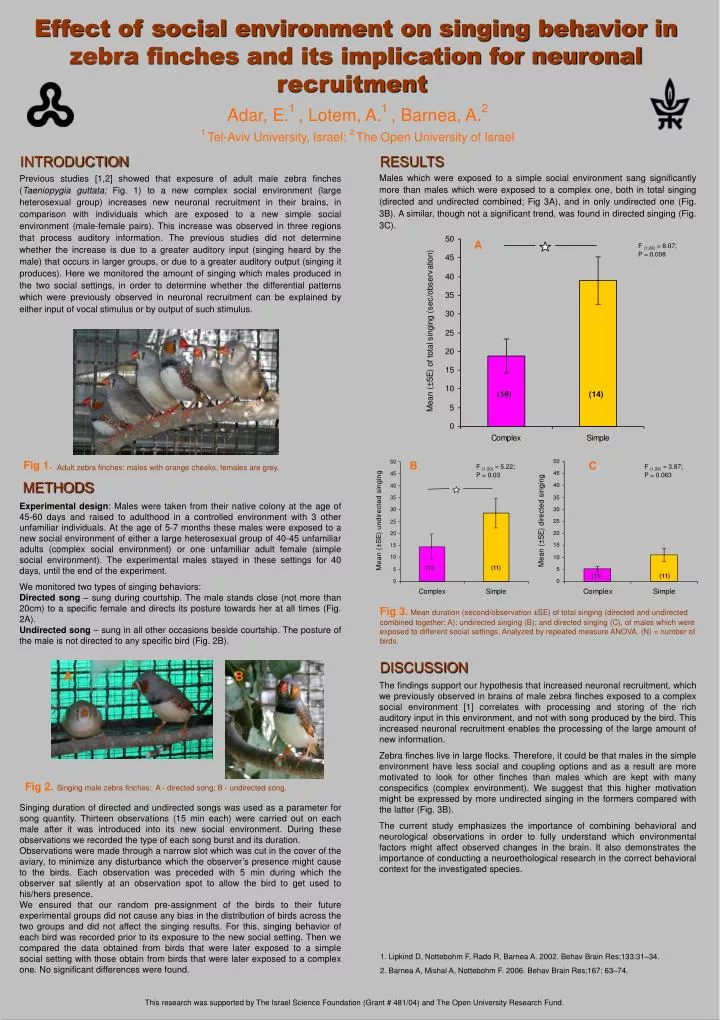
C. F (1,28) = 3.87; P = 0.063. A. B. Singing male zebra finches: A - directed song; B - undirected song. . Fig 2. Effect of social environment on singing behavior in zebra finches and its implication for neuronal recruitment . Adar, E. 1 , Lotem, A. 1 , Barnea, A. 2

E N D
C F (1,28) = 3.87; P = 0.063 A B Singing male zebra finches: A - directed song; B - undirected song. Fig 2. Effect of social environment on singing behavior in zebra finches and its implication for neuronal recruitment Adar, E.1 , Lotem, A.1 , Barnea, A.2 1 Tel-Aviv University, Israel; 2 The Open University ofIsrael INTRODUCTION RESULTS Males which were exposed to a simple social environment sang significantly more than males which were exposed to a complex one, both in total singing (directed and undirected combined; Fig 3A), and in only undirected one (Fig. 3B). A similar, though not a significant trend, was found in directed singing (Fig. 3C). Previous studies [1,2] showed that exposure of adult male zebra finches (Taeniopygia guttata; Fig. 1) to a new complex social environment (large heterosexual group) increases new neuronal recruitment in their brains, in comparison with individuals which are exposed to a new simple social environment (male-female pairs). This increase was observed in three regions that process auditory information. The previous studies did not determine whether the increase is due to a greater auditory input (singing heard by the male) that occurs in larger groups, or due to a greater auditory output (singing it produces). Here we monitored the amount of singing which males produced in the two social settings, in order to determine whether the differential patterns which were previously observed in neuronal recruitment can be explained by either input of vocal stimulus or by output of such stimulus. A F (1,28) = 8.07; P = 0.008 Fig 1. Adult zebra finches: males with orange cheeks, females are grey. B F (1,20) = 5.22; P = 0.03 METHODS Experimental design: Males were taken from their native colony at the age of 45-60 days and raised to adulthood in a controlled environment with 3 other unfamiliar individuals. At the age of 5-7 months these males were exposed to a new social environment of either a large heterosexual group of 40-45 unfamiliar adults (complex social environment) or one unfamiliar adult female (simple social environment). The experimental males stayed in these settings for 40 days, until the end of the experiment. We monitored two types of singing behaviors: Directed song – sung during courtship. The male stands close (not more than 20cm) to a specific female and directs its posture towards her at all times (Fig. 2A). Undirected song – sung in all other occasions beside courtship. The posture of the male is not directed to any specific bird (Fig. 2B). Fig 3. Mean duration (second/observation ±SE) of total singing (directed and undirected combined together; A); undirected singing (B); and directed singing (C), of males which were exposed to different social settings. Analyzed by repeated measure ANOVA. (N) = number of birds. DISCUSSION The findings support our hypothesis that increased neuronal recruitment, which we previously observed in brains of male zebra finches exposed to a complex social environment [1] correlates with processing and storing of the rich auditory input in this environment, and not with song produced by the bird. This increased neuronal recruitment enables the processing of the large amount of new information. Zebra finches live in large flocks. Therefore, it could be that males in the simple environment have less social and coupling options and as a result are more motivated to look for other finches than males which are kept with many conspecifics (complex environment). We suggest that this higher motivation might be expressed by more undirected singing in the formers compared with the latter (Fig. 3B). The current study emphasizes the importance of combining behavioral and neurological observations in order to fully understand which environmental factors might affect observed changes in the brain. It also demonstrates the importance of conducting a neuroethological research in the correct behavioral context for the investigated species. Singing duration of directed and undirected songs was used as a parameter for song quantity. Thirteen observations (15 min each) were carried out on each male after it was introduced into its new social environment. During these observations we recorded the type of each song burst and its duration. Observations were made through a narrow slot which was cut in the cover of the aviary, to minimize any disturbance which the observer’s presence might cause to the birds. Each observation was preceded with 5 min during which the observer sat silently at an observation spot to allow the bird to get used to his/hers presence. We ensured that our random pre-assignment of the birds to their future experimental groups did not cause any bias in the distribution of birds across the two groups and did not affect the singing results. For this, singing behavior of each bird was recorded prior to its exposure to the new social setting. Then we compared the data obtained from birds that were later exposed to a simple social setting with those obtain from birds that were later exposed to a complex one. No significant differences were found. • Lipkind D, Nottebohm F, Rado R, Barnea A. 2002. Behav Brain Res;133:31–34. • Barnea A, Mishal A, Nottebohm F. 2006. Behav Brain Res;167: 63–74. This research was supported by The Israel Science Foundation (Grant # 481/04) and The Open University Research Fund.