Download

1 / 58
590 likes | 739 Vues
Digestion and absorption of lipids and Fatty acid synthesis 脂类消化、吸收和脂肪酸合成. Deqiao Sheng PhD Biochemistry Department. Most lipids are ingested in the form of triacylglycerols. Dietary lipids are digested by pancreatic lipases .

E N D
Digestion and absorption of lipids and Fatty acid synthesis脂类消化、吸收和脂肪酸合成 Deqiao Sheng PhD Biochemistry Department
Most lipids are ingested in the form of triacylglycerols. Dietary lipids are digested by pancreatic lipases. Triacylglycerols in the intestinal lumen are incorporated into micelles formed with the aid of bile salts, amphipathic molecules synthesized from cholesterol in the liver and secreted from the gallbladder.
Most lipids are ingested in the form of triacylglycerols but must be degraded to fatty acids for absorption across the intestinal epithelium. Recall that lipids are not easily solubilized, yet they must be in order to be degraded. Triacylglycerols in the intestinal lumen are incorporated into micelles(微胶粒) formed with the aid of bile salts. Dietary Lipids Are Digested by Pancreatic Lipases
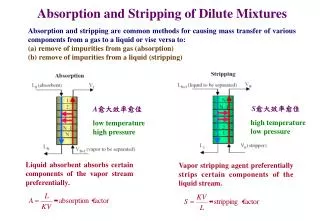
Most lipids in the diets of mammals are triacylglycerols with smaller amounts of phospholipids and cholesterol. The digestion of dietary lipids occurs mainly in the small intestine, where suspended fat particles are coated with bile salts. (Emulsification) Absorption
Bile salts are amphipathic cholesterol derivatives synthesized in the liver, collected in the gallbladder, and secreted into the lumen of the intestine. Bile salts play an important role in the absorption of dietary lipids. (Emulsification) Pancreatic lipase acts to degrade triacylglycerols in the fat particles. This lipase catalyzes hydrolysis at the C-1 and C-3 positions of a triacylglycerol, producing free fatty acids and a 2-monoacylglycerol.
pancreatic lipases Action of pancreatic lipase. Removal of the C-1 and C-3 acyl chains produces free fatty acids and a 2- monoacylglycerol.
A small protein called colipase(辅脂酶) helps bind the water-soluble lipase to the lipid substrates. Colipase also activates lipase by holding it in a conformation with an open active site. The initial products of fat hydrolysis are free fatty acids and monoacylglycerols. These molecules are transported to the intestinal wall in bile-salt micelles where they are absorbed by the cells lining the intestinal wall and the bile salts are released.
Fatty acids are converted to fatty acyl CoA molecules within the intestinal cells. Three of these molecules can combine with glycerol, or two with a monoacylglycerol, to form a triacylglycerol. These water-insoluble triglycerides combine with cholesterol and specific proteins to form chylomicrons for transport to other tissues.
Chylomicron Formation. Free fatty acids and monoacylglycerols are absorbed by intestinal epithelial cells. Triacylglycerols are resynthesized and packaged with other lipids and apoprotein B-48 to form chylomicrons, which are then released into the lymph system.
Triglyceridescombine with cholesterol and specific proteins to form chylomicrons(CM, 乳糜微粒) for transport to other tissues. The fate of dietary phospholipids is similar to that of triacylglycerols. Pancreatic phospholipases secreted into the intestine catalyze the hydrolysis of phospholipids, which aggregate in micelles. The major phospholipase in the pancreatic secretion is phospholipase A2, which catalyzes hydrolysis of the ester bond at C-2 of a glycerophospholipid to form a lysophosphoglyceride and a fatty acid.
Action of four phospholipases. Phospholipases A1, A2, C, and D can be used to dissect glycerophospholipid structure. Phospholipases catalyze the selective removal of fatty acids from C-1 or C-2 or convert glycerophospholipids to diacylglycerols or phosphatidates.
In the intestinal mucosal cells, the triacylglycerols are resynthesized from fatty acids and mono-acylglycerols and then packaged into lipoprotein (脂蛋白) transport particles called chylomicrons. These particles are composed mainly of triacylglycerols, with apoprotein B-48 as the main protein component. Protein constituents of lipoprotein particles are called apolipoproteins (载脂蛋白). Chylomicrons also function in the transport of fat-soluble vitamins and cholesterol. Dietary Lipids Are Transported in Chylomicrons
Triacylglycerols, cholesterol, and cholesteryl esterscannot be transported in blood or lymph as free molecules because they are insoluble in water. Instead, these lipids assemble with phospholipids and amphipathic lipid binding proteins to form spherical macromolecular particles known as lipoproteins. A lipoprotein has a hydrophobic core containing triacylglycerols and cholesteryl esters and a hydrophilic surface consisting of a layer of amphipathic molecules such as cholesterol, phospholipids, and proteins. Lipoproteins
The largest lipoproteins are chylomicrons, which deliver triacylglycerols and cholesterol from the intestine via the lymph and blood to tissues such as muscle (for oxidation) and adipose tissue (for storage) .
Fatty acids are synthesized by the repetitive addition of two-carbon units to the growing end of a hydrocarbon chain. Fatty acid biosynthesis and breakdown occur by different pathways, are catalyzed by different sets of enzymes, and take place in different parts of the cell.
ACP--acyl carrier protein An overview of fatty acid synthesis The first steps in the fatty acid synthesis pathway are the synthesis of acetyl ACP and malonyl ACP from acetyl CoA.
ACP--acyl carrier protein ACP Addition of two carbons to a growing fatty acyl chain: a four-step sequence.
Malonyl ACP is the main substrate for fatty acid biosynthesis. It is made in two steps, the first of which is the carboxylation of acetyl CoA in the cytosol to form malonyl CoA . The carboxylation reaction is catalyzed by the biotin- dependent enzyme acetyl-CoA carboxylase using a mechanism similar to the reaction catalyzed by pyruvate carboxylase . The ATP-dependent activation of HCO3- forms carboxybiotin. This reaction is followed by the transfer of activated CO2 to acetyl CoA, forming malonyl CoA. A. Synthesis of Malonyl ACP and Acetyl ACP
the first step The second step
The second step in the synthesis of malonyl ACP is the transfer of the malonyl moiety from coenzyme A to ACP. This reaction is catalyzed by malonyl CoA:ACP transacylase . A similar enzyme called acetyl CoA:ACP transacylase converts acetyl CoA to acetyl ACP.
The synthesis of long-chain fatty acids begins with the formation of a four-carbon unit attached to ACP. This molecule, called acetoacetyl ACP, is formed by condensation of a two-carbon substrate (acetyl CoA or acetyl ACP) and a three-carbon substrate (malonyl ACP) with the loss of CO2. The reaction is catalyzed by 3-ketoacyl ACP synthase (KAS). B. The Initiation Reaction of Fatty Acid Synthesis
Acetoacetyl ACP contains the smallest 3-ketoacyl moiety. In order to prepare for subsequent condensation reactions, this oxidized 3- ketoacyl moiety has to be reduced to an acyl form by the transfer of electrons (and protons) to the 3-carbon position. Three separate reactions are required. C. The Elongation Reactions of Fatty Acid Synthesis
In the first reduction the ketone is converted to an alcohol. The second step is the removal of water by a dehydratase producing a C=C double bond. Finally, a second reduction adds hydrogens to create the fully reduced acyl group. The final product of the reduction, dehydration, and reduction steps is an acyl ACP that is two carbons longer. This acyl ACP becomes the substrate for the elongation forms of 3-ketoacyl ACP synthase (KAS I and KAS II).
The end product of saturated fatty acid synthesis is 16- and 18-carbon fatty acids. Larger chain lengths cannot be accommodated in the binding site of the condensing enzyme. The elongation stage of fatty acid synthesis.
The thioesterase reaction results in release of free fatty acids but subsequent modifications of these fatty acids, and their incorporation into membrane lipids, require an activation step where they are converted to thioesters of coenzyme A in an ATP-dependent reaction catalyzed by acyl-CoA synthetase . D. Activation of Fatty Acids
The fatty acid synthase pathway cannot make fatty acids that are longer than 16 or 18 carbons (C16 or C18). Longer fatty acids are made by extending palmitoyl CoA or stearoyl CoA. The enzymes that catalyze such extensions are known as elongases and they use malonyl CoA (not malonyl ACP) as the source of the 2-carbon extension unit. Fatty Acid Extension and Desaturation
The complex is a dimer of two identical polypeptide monomers, 1 and 2, each consisting of seven enzyme activities and the acyl carrier protein (ACP). (Cys-SH, cysteinethiol.) The -SH of the 4′-phosphopantetheine of one monomer is in close proximity to the -SH of the cysteine residue of the ketoacyl synthase of the other monomer, suggesting a “head-to-tail” arrangement of the two monomers. The Fatty Acid Synthase Complex Is a Polypeptide Containing Seven Enzyme Activities
Though each monomer contains all the partial activities of the reaction sequence, the actual functional unit consists of one-half of one monomer interacting with the complementary half of the other. Thus, two acyl chains are produced simultaneously. The sequence of the enzymes in each monomer is based on Wakil.
acetyl malonyl Fatty acid synthase multienzyme complex By Salih Jawad Wakil was born in 1927 in Kerballa, Iraq
NADPH is involved as donor of reducing equivalents in both the reduction of the 3-ketoacyl and of the 2,3- unsaturated acyl derivatives. The oxidative reactions of the pentose phosphate pathway are the chief source of the hydrogen required for the reductive synthesis of fatty acids. The Main Source of NADPH for Lipogenesis Is the Pentose Phosphate Pathway
Palmitate, the principal product of the fatty acid synthase system in animal cells, is the precursor of other long-chain fatty acids . It may be lengthened to form stearate (18:0) or even longer saturated fatty acids by further additions of acetyl groups, through the action of fatty acid elongation systems present in the smooth endoplasmic reticulum and in mitochondria. Long-Chain Saturated Fatty Acids Are Synthesized from Palmitate
Routes of synthesis of other fatty acids. Palmitate is the precursor of stearate and longer-chain saturated fatty acids, as well as the monounsaturated acids palmitoleate and oleate. Mammals cannot convert oleate to linoleate or -linolenate (shaded pink), which are therefore required in the diet as essential fatty acids. Conversion of linoleate to other polyunsaturated fatty acids and eicosanoids is outlined. Unsaturated fatty acids are symbolized by indicating the number of carbons and the number and position of the double bonds.
Palmitate and stearate serve as precursors of the two most common monounsaturated fatty acids of animal tissues: palmitoleate, 16:1(Δ9), and oleate, 18:1(Δ9); both of these fatty acids have a single cis double bond between C-9 and C-10. Desaturation of Fatty Acids Requires a Mixed-Function Oxidase
The double bond is introduced into the fatty acid chain by an oxidative reaction catalyzed by fatty acyl–CoA desaturase , a mixed-function oxidase . Oxygenases, and Cytochrome P-450
Mammalian hepatocytes can readily introduce double bonds at the Δ9 position of fatty acids but cannot introduce additional double bonds between C-10 and the methyl-terminal end. Thus mammals cannot synthesize linoleate, 18:2(Δ 9,12), or -linolenate, 18:3(Δ 9,12,15). Because they are necessary precursors for the synthesis of other products, linoleate (亚油酸) and linolenate (亚麻酸) are essential fatty acids for mammals.
Synthesis of unsaturated FA unsaturated FA :软油酸、Oleate、linoleate、linolenate、arachidonic acid ( Essential FA ) Essential FA:required for the growth of mammals and they must be obtained from food. Including linoleate、linolenate, arachidonic acid
Overview ACP
Eicosanoids(廿碳) are a family of very potent biological signaling molecules that act as short-range messengers, affecting tissues near the cells that produce them. In response to hormonal or other stimuli, phospholipase A2, present in most types of mammalian cells, attacks membrane phospholipids, releasing arachidonate(花生四烯酸) from the middle carbon of glycerol. Eicosanoid Hormones Are Derived from Polyunsaturated Fatty Acids
Arachidonate, a 20:4 fatty acid derived from linoleate, is the major precursor of several classes of signal molecules: prostaglandins (PGs), prostacyclins, thromboxanes (TXs), and leukotrienes (LTs)
“Eicosanoid”(eicosa-, Greek for "twenty") is the collective term for oxygenated derivatives of three different 20-carbon essential fatty acids: Eicosapentaenoic acid (二十碳五烯酸 ,EPA), an ω-3 fatty acid with 5 double bonds; Arachidonic acid (AA), an ω-6 fatty acid, with 4 double bonds; Dihomo-gamma-linolenic acid (DGLA), an ω-6, with 3 double bonds.