
Biochemistry
E N D
Presentation Transcript
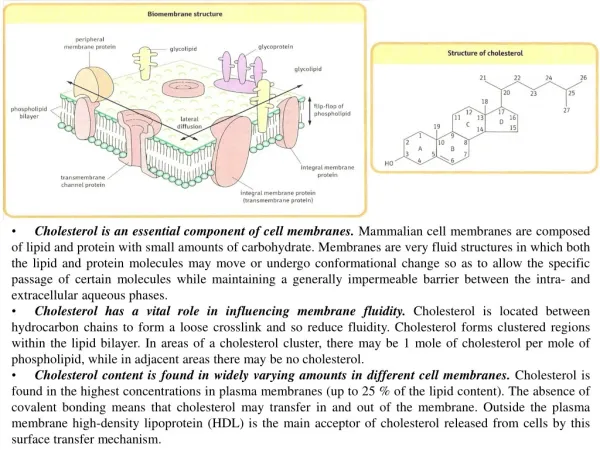
Cholesterol is an essential component of cell membranes. Mammalian cell membranes are composed of lipid and protein with small amounts of carbohydrate. Membranes are very fluid structures in which both the lipid and protein molecules may move or undergo conformational change so as to allow the specific passage of certain molecules while maintaining a generally impermeable barrier between the intra- and extracellular aqueous phases. • Cholesterol has a vital role in influencing membrane fluidity. Cholesterol is located between hydrocarbon chains to form a loose crosslink and so reduce fluidity. Cholesterol forms clustered regions within the lipid bilayer. In areas of a cholesterol cluster, there may be 1 mole of cholesterol per mole of phospholipid, while in adjacent areas there may be no cholesterol. • Cholesterol content is found in widely varying amounts in different cell membranes. Cholesterol is found in the highest concentrations in plasma membranes (up to 25 % of the lipid content). The absence of covalent bonding means that cholesterol may transfer in and out of the membrane. Outside the plasma membrane high-density lipoprotein (HDL) is the main acceptor of cholesterol released from cells by this surface transfer mechanism.
BILE ACIDS Quantitatively the most important metabolic products of cholesterol are the bile acids. In man there are four main bile acids. These bile acids all have 24 carbon atoms with the terminal three carbon atoms of the cholesterol side chain removed during synthesis Biosynthesis of bile acids occurs within the liver, parenchymal cells. Biosynthesis occurs within the liver parenchymal cells to produce cholic and chenodeoxycholic acids. Prior to secretion these primary bile acids are conjugated through the carboxyl group forming amide linkages with either glycine or taurine. In man there is a 3:1 ratio in favor of glycine conjugates. These compounds are secreted from the liver via larger bile ducts, either directly into the duodenum or for storage in the gall bladder. Deoxycholic and lithocholic acids are secondary bile acids formed within the intestine through the action of bacterial enzymes on the primary bile acids.
STEROID HORMONES • Cholesterol is the precursor of all of the steroid hormones. The biosynthesis of the steroid hormones occurs within the adrenal cortex, the testis in man and the ovary in woman. In pathologic situations, such as a defect in steroidogenesis or a steroid-secreting tumor, a very abnormal pattern of steroid secretion may be observed. • Most of the enzymes involved in converting cholesterol into steroid hormones are cytochrome P450 proteins that require oxygen and NADPH. • Biosynthesis of the corticosteroids. The cellular substructure of the adrenal cortex is arranged in three different layers. The inner two layers are responsible for the synthesis of cortisol, the main glucocorticoid. The outer layer is responsible for the synthesis of aldosterone, the main mineralocorticoid. Although many of the steps are similar they are controled by very different mechanisms and this has led to the suggestion that the adrenal cortex may be considered as two separate endocrine organs. • The biosynthesis of cortisol depends on stimulation by pituitary adrenocorticotropic hormone (ACTH). • The main stimulus to the synthesis of aldosterone is angiotensin II produced as a result of the enzymes of the renin-angiotensin system. Potassium is an important secondary stimulus. • Biosynthesis of the androgens. The conversion of corticosteroids into androgens requires the P450c17 17-20 lyase/desmolase enzyme. This enzyme is abundant in the Leydig cells of the testis and in the granulosa cells of the ovary. In these cases, however, the stimuli to the rate-limiting cholesterol side-chain cleavage step are luteinizing hormone (LH) and follicle stimulating hormone (FSH), respectively. • Biosynthesis of the estrogens. The conversion of androgens into estrogens involves action of P450c19 19-aromatase. This enzyme is found most abundantly in the granulosa cells of the ovary although it enzyme in adipose tissue can also convert some testosterone into estradiol.
The liver plays a central role in the regulation of the body’s cholesterol homeostasis. Cholesterol enters the liver’s cholesterol pool from a number of sources including dietary cholesterol, as well as cholesterol synthesized denovo by extrahepatic tissues and by the liver itself. Cholesterol is eliminated from the liver as unmodified cholesterol in the bile, or it can be converted to bile salts that are secreted into the intestinal lumen. In humans, the balance between cholesterol influx and efflux is not precise, resulting in a gradual deposition of cholesterol in the tissues, particularly in the endothelial linings of blood vessels. This is a potentially life-threatening occurrence when the lipid deposition leads to plaque formation, causing the narrowing of blood vessels (atherosclerosis) and increased risk of cardio-, cerebro- and peripheral vascular disease.
SYNTHESIS OF 3-HYDROXY-3METHYLGLUTARYL (HMG) COA SYNTHESIS OF MEVALONATE The reduction of HMG CoA to mevalonate, is catalyzed by HMG CoA reductase, and is the rate-limiting and key regulated step in cholesterol synthesis. It occurs in the cytosol, uses two molecules of NADPH as the reducing agent, and releases CoA, making the reaction irreversible. The first two reactions in the cholesterol synthetic pathway. Result in the production of HMG CoA. Liver parenchymal cells contain two isoenzymes of HMG CoA synthase. The cytosolic enzyme participates in cholesterol synthesis, whereas the mitochondrial enzyme functions in the pathway for ketone body synthesis.
REGULATION OF CHOLESTEROL SYNTHESIS HMG CoA reductase, the rate-limiting enzyme, is subject to different kinds of metabolic control. • Sterol-dependent regulation of gene expression • Sterol-accelerated enzyme degradation • Sterol-independent phosphorylation/dephosphorylation: Through the actions of adenosine monophosphate (AMP)-activated protein kinase and a phosphoprotein phosphatase. The phosphorylated form of the enzyme is inactive, whereas the dephosphorylated form is active. • Hormonal regulation: An increase in insulin and thyroxine favors up-regulation of the expression of the gene for HMG CoA reductase. Glucagon and the glucocorticoids have the opposite effect. • Inhibition by drugs: The statin drugs (fluvastatin, lovastatin, pravastatin, etc.) are structural analogs of HMG CoA, and are reversible, competitive inhibitors of HMG CoA reductase. They are used to decrease plasma cholesterol levels in patients with hypercholesterolemia.
PLASMA LIPOPROTEINS The plasma lipoprotein are spherical macromolecular complexes of lipids and specific proteins. Lipoproteins function both to keep their component lipids soluble as they transport them in the plasma and to provide an efficient mechanism for transporting their lipid contents to (and from) the tissues. Apolipoprotein: The apolipoproteins associated with lipoprotein particles are providing recognition sites for cell-surface receptors, and serving as activators or coenzymes for enzymes involved in lipoprotein metabolism.
METABOLISM OF CHYLOMICRONS • Synthesis of apolipoproteins: Apolipoprotein B-48 is unique to chylomicrons. • Assembly of chylomicrons: Assembly of the apolipoproteins and lipid into chylomicrons requires microsomal triacylglycerol transfer protein which loads apo B-48 with lipid. • Modification of nascent chylomicron particles: The particle released by the intestinal mucosal cell is called a “nascent” chylomicron because it is functionally incomplete. When it reaches the plasma, the particle is rapidly modified, receiving apolipoprotein E and C. The latter includes apo C-II, which is necessary for the activation of lipoprotein lipase. The source of these apolipoproteins is circulating HDL. • Degradation of triacylglycerol by lipoprotein lipase. • Formation of chylomicron remnants: As the chylomicron circulates and more than 90% of the triacylglycerol in its core is degraded by lipoprotein lipase, the particle decreases in size and increases in density. In addition, the C apoproteins are returned to HDL. The remaining particle, called a “remnant”, is rapidly removed from the circulation by the liver, whose cell membranes contain lipoprotein receptors that recognize apo E.
METABOLISM OF VLDL VLDLs are produced in the liver. They are composed predominantly of endogenous triacylglycerol (approximately 60%), and their function is to carry this lipid from the liver to the peripheral tissues. • Release of VLDL: VLDL are secreted directly into the blood by the liver as nascent VLDL particles containing apo B-100. They must obtain apo C-11 and apo E from circulating HDL. • Modification of circulating VLDL: As VLDL pass through the circulation VLDL decrease in size and become denser, surface components, including the C and E apoproteins, are returned to HDL, but the particles retain apo B-100. • Production of LDL from VLDL in the plasma.
METABOLISM OF LDL LDL particles contain much less triacylglycerol than their VLDL and have a high concentration of cholesterol and cholesterol esters. • Receptor-mediated endocytosis: The primary function of LDL particles is to provide cholesterol to the peripheral tissues. They do so by binding to cell surface membrane LDL receptors that recognize apo B-100. • 1. LDL receptors are negatively charged glycoproteins that are clustered in pits on cell membranes. The cytosolic side of the pit is coated with the protein clathrin, which stabilizes the shape of the pit. • 2. After binding, the LDL-receptor complex is internalized by endocytosis. A deficiency of functional LDL receptors causes type II hyperlipidemia and premature atherosclerosis. • 3. The vesicle containing LDL loses its clathrin coat and fuses with other similar vesicles, forming larger vesicles called endosomes. • 4. The pH of the endosome falls . The receptors then migrate to one side of the endosome, whereas the LDLs stay free within the lumen of the vesicle. • 5. The receptors can be recycled, whereas the lipoprotein remnants in the vesicle are transferred to lysosomes and degraded by lysosomal acid hydrolases, releasing free cholesterol, amino acids, fatty acids, and phospholipids. These compounds can be reutilized by the cell. • Effect of endocytosed cholesterol on cellular cholesterol homeostasis: The chylomicron remnant-, IDL-, and LDL-derived cholesterol affects cellular cholesterol content in several ways. • 1. HMG CoA reductase is inhibited by high cholesterol denovo cholesterol synthesis decreases. • 2. Synthesis of new LDL receptor protein is reduced by decreasing the expression of the LDL receptor gene.
LIPOPROTEIN RECEPTORS The LDL receptor allows cells to acquire cholesterol from outside the cells. The receptor gene is located on chromosome 19; the mature receptor protein is 839 amino acids long and spans the cell membrane. The receptor was discovered by Goldstein and Brown, who jointly received the Nobel Prize for their work. The LDL receptor is also known as the apoB/E receptor, because it has an affinity to two apoproteins: apoE and apoB100. ApoE binds to LDL receptor with a higher affinity than apoB. ApoB48 does not bind to the LDL receptor. The scavenger receptor:Scavenger receptors are membrane receptors with a relatively broad specificity. They are present on the phagocytic cells such as macrophages. While intact LDL does not interact with the scavenger receptor, chemically modified (acetylated or oxidized LDL) is readily bound.
METABOLISM OF HDL HDL particles are formed in blood by the addition of lipid to apo A-1, an apolipoprotein made by the liver and intestine and secreted into blood. • HDL is a reservoir of apolipoproteins: HDL particles serve as a circulating reservoir of apo C-II and apo E. • HDL uptake of unesterified cholesterol: Nascent HDL are disk-shaped particles containing primarily phospholipid and apolipoproteins A, C, and E. They take up cholesterol from non-hepatic (peripheral) tissues and return it to the liver as cholesteryl esters. • Esterification of cholesterol: When cholesterol is taken up by HDL, it is immediately esterified by the plasma enzyme lecithin:cholesterol acyltransferase (LCAT). This enzyme is synthesized by the liver. LCAT binds to nascent HDL, and is activated by apo A-I. LCAT transfers the fatty acid from carbon 2 of phosphatidylcholine to cholesterol.
REVERSE CHOLESTEROL TRANSPORT The selective transfer of cholesterol from peripheral cells to HDL, and from HDL to the liver for bile acid synthesis or disposal via the bile, and to steroidogenic cells for hormone synthesis, is a key component of cholesterol homeostasis. This is, in part, the basis for the inverse relationship seen between plasma HDL concentration and atherosclerosis, and for HDL’s designation as the “good” cholesterol carrier. Reverse cholesterol transport involves 1. Efflux of cholesterol from peripheral cells to HDL 2. Esterification of cholesterol by LCAT 3. Binding of the cholesteryl ester-rich HDL to liver and steroidogenic cells 4. The selective transfer of the cholesteryl esters into these cells 5. The release of lipid-depleted HDL It is not yet clear as to whether the HDL particle itself is taken up, the cholesteryl esters extracted, and the lipid-poor HDL released back into the blood, or if there is selective uptake of the cholesteryl ester alone.
PRIMARY DISORDERS OF PLASMA LIPOPROTEINS (DYSLIPOPROTEINEMIAS)