Download

1 / 71
790 likes | 1.8k Vues
Stability of mRNA. Structure features of eukaryotic mRNA untranslated regions (UTR) Regulation of mRNA stability in mammalian cells Bioinformatic analysis of UTR functional characterization. Significance of mRNA Stability.

E N D
Stability of mRNA • Structure features of eukaryotic mRNA untranslated regions (UTR) • Regulation of mRNA stability in mammalian cells • Bioinformatic analysis of UTR functional characterization
Significance of mRNA Stability Controlling the rate at which the mRNA decays can regulate the levels of cellular messenger RNA transcripts. Because decay rates affect the expression of specific genes, they provide a cell with flexibility in effecting rapid change.
mRNA abundance is determined by balancing transcription and RNA decay. mRNA stability can be rapidly modulated to alter the expression of specific genes thereby providing flexibility in affecting changes in patterns of protein synthesis.
In eukaryotes: the 3’ poly(A) tail confers stability. In bacteria: a hairpin structure in mRNA with r-independent terminator confers stability.
Moreover, many clinically relevant mRNAs--including several encoding cytokines, growth factors and proto-oncogenes--are regulated by differential RNA stability.
mRNA Decay and Translation The intimate relationship between mRNA decay and translation is further indicated by the ability of translation-initiation factors (eIF) and proteins (PAB) that bind the poly(A) tail to protect the mRNA from degradation. Moreover, evidence shows that inhibiting translation elongation promotes mRNA stabilization.
What is the rate-limiting step in mRNA degradation? An evolutionarily conserved mRNA-degradation pathway is initiated by the removal of the 3’- poly(A) tail. This disrupts the translation initiation complex and provides degradative enzymes with access to the 5’ cap and remaining RNA body.
What are the sequence elements and factors that control the half-lives of mRNAs?
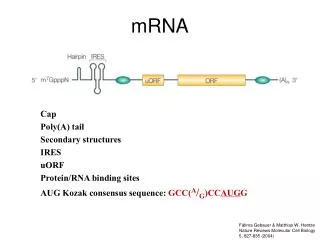
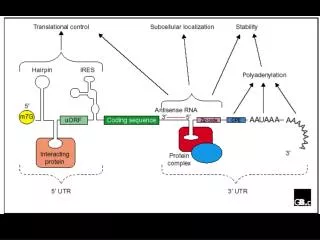
General Structure of a Eukaryotic mRNA illustrating some post-transcriptional regulatory elements for gene expression and their activity. 5’UTR mediated regulation may involve: the 7-methyl-guanine (cap); hairpin-like secondary structure; RNA-protein interactions; upstream open reading frames (uORFs); internal ribosome entry sites (IRES). 3’UTR mediated regulation may involve: antisense RNA interactions; RNA-protein interactions, involving also multiprotein complexes; cytoplasmic polyadenylation elements (CPE); poly(A) tail and variation of its size.
Elements in the 5’ UTR 7-methylguanosine cap
Caps provide at least four functions: • Protection of mRNA from degradation (see example) • Enhancement of mRNA’s translatability • Transport of mRNA out of nucleus • Proper splicing of pre-mRNA
Furuichi & colleagues, 1977 Labeled reovirus RNAs w/ capped (green), blocked (blue) or decapped (red) 5’-end a) glycerol gradient centrifugation (ggc) b) incubation in X. oocyte, 8 h > ggc c) decapped & deblocked RNAs (b-elimination yielding pppGm & pppG) + incubation in X. oocyte, 8 h > ggc d) as (b) but incubation in wheat germ extract w/o w/
Elements in the 3’ UTR • AU-rich element (ARE) and proteins that bind AREs • Iron-responsive element (IRE) and iron regulatory protein (IRP) • Cell cycle-regulated histone mRNA stem-loop determinant (SL/SLBP) • Cytoplasmic polyadenylation element (CPE)……
Transferrin receptor mRNAion-responsive element & IRP PNAS 93: 8179-8182, 1996
Histone mRNA stem loop determinant & SLBP Gene 239: 1-14, 1999
assembly CPSF: blue CstF: brown CF I & II: grey cleavage stimulated by pol II CTD CstF & CFs leaves PAP (orange) enters Cytoplasmic polyadenylation elements (CPE) & binding proteins oligo(A) synthesis aided by CPSF poly(A) synthesis aided by PAB II (yellow)
Darnell & colleagues, 1973 Poly(A): nuclear form 210 nt, cytoplasmic form 190 nt > poly(A) undergoes considerable shortening in the cytoplasm Nuclear cytoplasmic 5S rRNA
Cytoplasmic polyadenylation of maternal mRNAs The best studied cases are those that occur during oocyte maturation Maturation-specific polyadenylation of Xenopusmaternal mRNAs in the cytoplasm depends on two sequence motifs: The AAUAAA motif & an upstream motif with UUUUUAU or a closely related sequence
Wickens & colleagues, 1989 inject labeled RNAs into X. oocyte cytoplasm > stimulate maturation w/ progesterone > isolate RNAs & electrophoresis Lack UUUUUAU • maturation-specific (i.e., D7) RNAs, containing UUUUUAU, • was polyadenylated P: pregesterone A: separation by oligo(dT)
AAUAAA is also required for cytoplasmic polyadenylation • this motif is required for both nuclear & cytoplasmic polyadenylation SV40 RNA
Poly(A) is not just shortened in the cytoplasm; it turns over! RNases tear it down, and poly(A) polymerase builds it back up When the poly(A) is gone, the mRNA is slated for destruction
Stability of mRNA • Structure features of eukaryotic mRNA untranslated regions (UTR) • Regulation of mRNA stability in mammalian cells • Bioinformatic analysis of UTR functional characterization
mRNA Decay Pathways in Mammalian Cells • Deadenylation-dependent pathways • Deadenylation-independent pathways: 1. endoribonucleolytic decay 2. nonsense-mediated decay (NMD) • RNAi-dependent pathway
Deadenylation-dependent mRNA Decay When mRNA processing is complete, the mRNA bears a 5' cap structure and 3' poly(A) tail that protect the message from exonucleolytic decay. The first step in the decay of most wild-type mRNAs is shortening of the poly(A) tail by a deadenylase (blue). Once poly(A) shortening is complete, the 5' 7-methylguanosine cap is rapidly removed and the rest of the mRNA is attacked by 5' and 3' exonucleases (green and pink, respectively).
The turnover of mRNAs is also regulated by cis-acting elements that either promote or inhibit their decay. The most prevalent is the AU-rich element (ARE), found in the 3’-untranslated region (3’ UTR) of mRNAs encoding many important growth control proteins.
The deadenylase as an inhibitor of translation initiation and decapping During translation, the mRNA is thought to be circularized by its interaction with the translation-initiation factors eIF4E (4E), eIF4G (4G) and the poly(A)-binding protein (PABP). This conformation protects the 3' and 5' ends of the mRNA from attack by the deadenylase and decapping enzymes. The deadenylase can somehow invade this closed loop and interact with the cap while simultaneously removing the poly(A) tail. The interaction of poly(A) ribonuclease (PARN) with the cap perpetuates the closed loop and thereby blocks both translation initiation and decapping.
When poly(A) shortening is complete, PARN dissociates, allowing the decapping enzyme to hydrolyse the 5' cap of the message. Meaning that hydrolysis the 5' cap of mRNA depends on the completion of poly(A) shortening by PARN.
In addition, ARE-binding proteins affect mRNA stability, translation and subcellular localization. Other elements found in the 5’ UTR and coding regions also modulate transcript stability.
Signalling pathways also affect mRNA stability Several signaling pathways are implicated in triggering changes in stability of specific mRNAs. e.g., interleukin-2 mRNA, which is stabilized by the c-Jun amino-terminal kinase (JNK) signaling pathway through JNK-responsive elements in its 5’ UTR.
AU-rich element: The most prevalent cis-acting element Class I AREs contain 1 to 3 scattered copies of the pentanucleotide AUUUA embedded within a U-rich region, and are found in the c-Fos and c-Myc mRNAs. Class II AREs contain multiple overlapping copies of the AUUUA motif, and are found in cytokine mRNAs. Class III AREs, such as the one in c-Jun mRNA, lack the hallmark AUUUA pentanucleotide but present U-rich sequences.
Model for how the ARE mediates stability and instability Interaction of the ARE with a destabilizing factor, such as AUF1 [to c-myc mRNA], might promote rapid deadenylation by reducing the affinity of the poly(A) binding protein (PABP) for the poly(A) tail. Conversely, stabilizing factors, such as HuR [to VEGF mRNA in response to hypoxia], might enhance binding of the PABP to the poly(A) tail, thus blocking deadenylation.
Some other ARE-BPs have been proven to possess destabilizing activity on ARE-RNAs: TTP, BRF1, KSRP AUF1 has a dual role in ARE-mediated mRNA decay, functioning either as a destabilizing or a stabilizing factor depending on the cell type.
mRNA Decay Pathways in Mammalian Cells • Deadenylation-dependent pathways • Deadenylation-independent pathways: 1. endoribonucleolytic decay 2. nonsense-mediated decay (NMD) • RNAi-dependent pathway
Endoribonucleolytic Decay There are a few messenger RNAs that degrade by a minor pathway known as endoribonucleolytic decay. Endoribonucleases recognize specific sequence elements within the transcript and cleave the mRNA internally. The cleavage event generates free 3' and 5' ends that are easily accessible to exonucleases and the products of the cleavage reaction are therefore rapidly degraded. In contrast, stabilizer protein may block the binding of endoribonuclease.
Interestingly, several mRNAs that are degraded by endoribonucleolytic decay also interact with stabilizer proteins that block access of the endoribonuclease to its cleavage site. e.g., an endonuclease from Xenopus laevis hepatocytes, PMR1, can cleave the vitellogenin mRNA but its action is prevented by binding of the vigilin protein to a site that overlaps the PMR1 cleavage site. Similarly, the a-globin mRNA is cleaved at a site in its 3' UTR by an erythroid-enriched endonuclease. In this case, cleavage is inhibited by binding of the a-CP complex of proteins to an overlapping sequence.
A strong link between translation and RNA turnover is also shown by nonsense-mediated decay (NMD), which ensures that mRNAs containing premature stop codons are degraded. (evidence from yeast studies)
NMD Pathways NMD prevents the accumulation of aberrant transcripts and truncated proteins by ensuring rapid decay of the mRNAs. Targets: transcripts that harbor nonsense codons, unspliced introns or extended 3’UTRs.
In the yeast S. cerevasiae, NMD involves deadenylation-independent decapping followed by 5’-3’ degradation of mRNA. • In mammalian cells, the sequence of degradation steps involved in NMD is unknown.
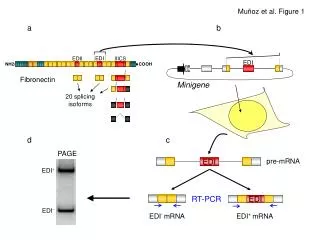
NMD of alternatively spliced mRNA (Bioinformatics 19: 118-121, 2003)
Translation OK Remaining EJCs trigger NMD (Bioinformatics 19: 118-121, 2003)
Regulated mRNA Stability • Regulation in response to developmental/differentiation cues • Regulation in response to hormonal regulation • Regulation in response to diurnal variation • Regulation in response to stress