Download

1 / 29
290 likes | 445 Vues
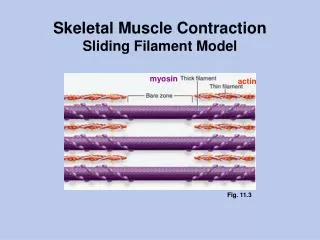
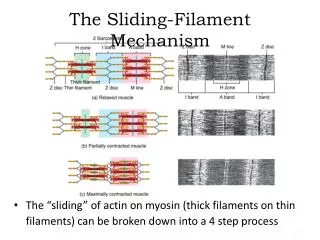
Sliding filament theory AF Huxley and R Niedergerke, 1954. Tropomyosin and troponin regulate the interaction between actin and myosin proteins of thick and thin filaments During contraction, cross-bridges attach between actin and myosin

E N D
Sliding filament theory AF Huxley and R Niedergerke, 1954 Tropomyosin and troponin regulate the interaction between actin and myosin proteins of thick and thin filaments During contraction, cross-bridges attach between actin and myosin Two filaments slide over each other when energy is provided by the hydrolysis of ATP
Neuromuscular Bases of Contraction Skeletal muscle contracts only after stimulation from a motor neuron Normally, each motor neuron branches several times and stimulates a few to several hundred muscle fibers
Motor Unit • Motor neuron, (cell, etc.) • Muscle fibers it innervates
Site: neuromuscular junction, motor end plate, myoneural junction
Contraction begins • Initiation of the action potential by the motor neuron • Transmission of the AP across the motor end plate to the muscle fiber
when AP reaches neuromuscular junction • ~ 200-300 vesicles of acetycholine (ACH, neurotransmitter) are released into the gap between the motor neuron and the motor end plate (cleft)
ACH diffused across the gap and reacts with receptor molecules in the sarcolemma • Reaction causes an increase in permeability to sodium ions, resulting in depolarization of the sarcolemma or end-plate potential • If end-plate potential is large enough to exceed a threshold (depending on skeletal muscle type), the nerve impulse will be successfully transformed into a muscle impulse
The impulse travels in all directions over the muscle membrane when being transmitted • Deep into the fiber through the transverse tubules (t-tubules)
ACH is released from the vesicles and diffuses across the cleft • binds with receptor, increasing the permeability of the sarcolemma to sodium ions • depolarizing the sarcolemma (end-plate potential) • Impulse travels down t-tubules, all over membrane to transverse tubules
As the AP is transmitted throughout the fiber, the membranes of the cisterane in the SR become more permeable to Ca++ and the stored ions diffuse into the sarcoplasm of the fiber • Once the Ca++ concentration is high enough, (100X increase, 10-5 M), the Ca++ binds with the TnC molecule
Binding of Ca++ to the TnC causes a positional change of the Tn, which also effects the positioning of the tropomyosin, moving it deeper into the groove between the two actin strands
TnC • Two different isoforms • One specific to fast muscle • One specific to slow muscle • Fast contain two binding sites for Ca++ • Site I and site II • Slow have only one binding site • both sites must be filled to trigger contraction
Conformational Change • there is a conformational change with binding that exposes a hydrophobic cavity (the TnI binding site). • Alters the interaction between TnI and TnC • Instead of TnI binding to actin, it preferentially switches to binding domain on TnC, allowing actin and myosin to interact.
Slow and Cardiac Muscle • Slow skeletal muscle has no site I • Slow and cardiac muscle are activated by one, not two calcium ions by the TnC isoforms subunit. • Therefore, contraction frequency, power output, and strength are typically down regulated • all of the characteristics of these subunits and their role in contraction are not yet clear • (e.g., Tn complex may attach the tropomyosin to the actin) do know that each subunit plays a role in the contractile process
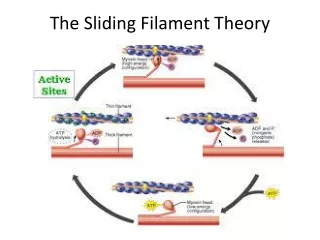
Actin binding sites • Uncovered, actin and myosin can interact • ATPase activity of myosin head immediately hydrolyzes ATP • Conformation of the head is that it extends perpendicularly towards the actin filament at this time
The products of the hydrolysis (ADP and Pi) remain bound to the head, which is now “energized” with the energy released from the reaction • Myosin head now interacts with the binding sites on the actin filament • When actin and myosin bind, forming an actomyosin complex, the stored energy is released
This release of energy alters the position of the myosin head and produces force through the cross-bridge movement • the head tilts toward the arm of the cross bridge, providing the power stroke for pulling the actin
the energy activating the power stroke is the energy already stored, like a cocked spring, by the conformational change in the head when the ATP molecule cleaved • when the head attaches to the active site, there are changes in the intramolecular forces between the head and the arm of the cross-bridge
this alignment of forces causes the head to tilt toward the arm and to drag the actin filament along with it • this tilt of the head is called the power stroke, and causes the release of ADP and Pi • immediately after tilting, the head automatically breaks away from the active site, binding an ATP
the head returns to the perpendicular position • once head is detached, a new molecule of ATP is hydrolyzed by the myosin ATPase, energizing the head again so cycle can repeat • In this new position, it binds with a new actin binding site
heads of cross-bridges bend back and forth, step by step, walking along the actin filaments toward the center of the myosin • each myosin acts independent of each other, each attaching and pulling in a continuous but random cycle • the greater the number of cross-bridges in contact with actin at any given time, the greater the theoretical force of contraction
because of the orientation of the actin and myosin, the actin filaments move towards each other, the Z lines move closer together and the H zone disappears • this process will continue until the Z membrane is pulled against the myosin filament or until the load on the muscle becomes too great for further pulling to occur (assuming muscle stimulation is still occurring)
Single Contraction Cycle • Contraction cycle of myosin cross-bridges of a muscle shortens a muscle by 1% of its resting length • consequently, the contraction cycle must be repeated over and over to significantly shorten the whole muscle
Fenn Effect • When a new ATP attaches to a myosin head, the cross-bridge can detach from the actin and the greater amount of work performed by the muscle, the greater amount of ATP which is cleaved
ACH • at same time contraction is occurring, ACH that stimulated the contraction is being rapidly decomposed by the action of cholinesterase (enzyme present at the myoneural junction within the membranes of the motor end plate) • rapid removal of ACH insures that a single nerve impulse will not cause a continued stimulation of the muslce
Impulse Duration • Usual duration of an impulse to skeletal muscle is about 20 milliseconds • in order for contraction to continue, there must be continual stimulation of the muscle fiber • the signal to stop contraction is the absence of a nerve impulse at the junction
AP stops • continually active calcium pump located in the walls of the SR pumps the calcium ions out of the sarcoplasm and back into the SR via the fenestrated collar, and then the calcium diffuses back into the cisternae
this lowers the concentration of calcium, removing it from the TnC, the Tn/tropomyosin complex returns to its original conformation and the active sites are covered • fiber returns to its relaxed position