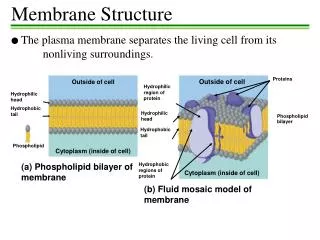
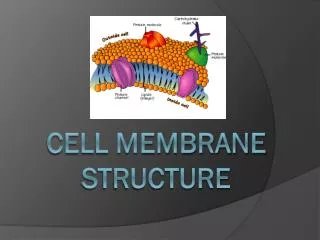
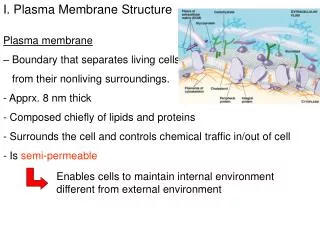
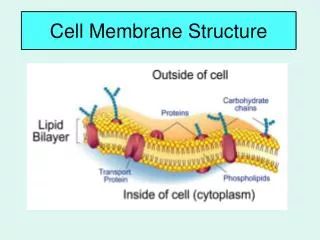
STRUCTURE DE LA MEMBRANE
STRUCTURE DE LA MEMBRANE. II - LES PROTÉINES MEMBRANAIRES. Porteuses de la plupart des fonctions des membranes Variable en fonction du type de cellule myéline : protéines 25 % de la masse membrane interne des mitochondries : protéines 75 % de la masse


STRUCTURE DE LA MEMBRANE
E N D
Presentation Transcript
II - LES PROTÉINES MEMBRANAIRES • Porteuses de la plupart des fonctions des membranes • Variable en fonction du type de cellule • myéline : protéines 25 % de la masse • membrane interne des mitochondries : protéines 75 % de la masse • en général : protéines 50 % de la masse • 50 molécules de lipide pour une molécule de protéine • Souvent glycosylées
Modes d'insertion des protéines dans la double couche lipidique (1/2) Fig 10-17 (1à4) Simple hélice Cylindre Multiple hélice
Modes d'insertion des protéines dans la double couche lipidique (2/2) Fig 10-17 ( 5 à 8) Phosphatidyl inositol Au départ protéine transmembranaire à un passage dans le RE Chaine d'acide gras ou groupe prényl. Au départ protéine cytosolique soluble
Protéines liées aux lipides Exemple 5 • Synthèse de la protéine dans le cytosol comme une protéine soluble • Fixation d'un lipide par liaison covalente • Transport vers la membrane
Fig 10-18 Exemple 5 : attachement d'une protéine membranaire par une chaîne d'acide gras ou un groupement prényl
Exemple 5 : attachement d'une protéine membranaire par une chaîne d'acide gras ou un groupement prényl • Aide à localiser une protéine soluble dans la membrane • Peut être transitoire
Exemple 6 : protéines liées aux lipides • Synthèse de la protéine dans le RE comme une protéine transmembranaire à un passage • Dans le RE le segment transmembranaire de la protéine est clivé • Un GlycosylPhosphatidylnositol (GPI) est ajouté • La protéine se retrouve liée à la face non cytosolique de la membrane • Facile à reconnaître grâce à une phospholipase C spécifique du phosphatidylInositol
Modes d'insertion des protéines dans la double couche lipidique (Rappel) Fig 10-17 ( 5 à 8)
Exemples 7 et 8 : protéines périphériques • Liaisons non covalentes avec d'autres protéines de la membrane • Peuvent être libérées par les procédés d'extraction doux (force ionique ou pH)
Protéines intégrales • = protéines intrinsèques • Nécessitent des procédés énergiques pour être libérées
Protéines membranaires • Transmembranaires • agissent des deux côtés de la membrane • transport de molécules • comprennent les récepteurs • reconnaissance extra cellulaire • message intra cellulaire • Non transmembranaires • associées aux lipides et/ou protéines d'une seule face de la membrane • eg : médiation chimique intra cellulaire (moitié cytosolique de la membrane)
Protéines transmembranaires • Toujours orientation unique dans la membrane • Dû à la synthèse dans le réticulum endoplasmique • Presque toujours en hélice • Ou cylindre
Hélice Fig 10-19
Fig 10-20 (AB) Prédiction de la localisation d'hélice d'une protéine par profil d'hydrophobicité
Fig 10-20 C Proportion prévisible de protéines membranaires dans le génome
Cylindres • Forment une structure rigide • facile à cristalliser ( hélice ) • Le nombre de plis peut varier de 8 à 22
Fig 10-21 (1de2) • Cylindres E. coli E. coli Récepteur à un virus bactérien Lipase
L'intérieur est comblé par un domaine globulaire de protéine qui contient un site de fixation du fer Fig 10-21(2de2) • Cylindres Rhodobacter capsulatus E. coli Transporteur d'ions
Les Cylindres • Abondants dans la membrane externe des mitochondries • Forment des pores (eg : porine) • Souvent boucles qui font saillie dans la lumière • Parfois porines spécifiques (maltoporines) • Surtout membranes externes des bactéries, mitochondries, chloroplastes • Beaucoup plus rares que hélices chez les eucaryotes • Plus rigides que l'hélice
Forrest,LR2000(Fig1) Review Membrane simulations: bigger and better? Simulations d'une hélice de protéine transmembranaire dans une bicouche lipidique. Simulations de (a) 18-mer, (b) 26-mer and (c) 34-mer modèles de l'hélice de la protéine transmembranaire M2 du virus influenza A dans une bicouche lipidique
Forrest,LR2000(Fig2) Review Membrane simulations: bigger and better? Simulations de canaux ioniques dans une bicouche lipidique. (a) Hélice M25 (b) Canal potassique KcsA
Fig 10-22 • Protéine transmembranaire à un passage glycosylée • Glycosylation dans le RE et Golgi • La face glycosylée est la face non cytosolique • Ponts S—S sur la face non cytosolique
C - Détergents • Petites molécules amphiphiles • Permettent de solubiliser les protéines transmembranaires • Extrémité polaire chargée (ionique) : sodium dodécyl sulfate (SDS) • Extrémité polaire non chargée (non ionique) : triton
Fig 10-23 • Micelle de détergent dans l'eau
Fig 10-24 • Solubilisation de protéines membranaires par un détergent léger: il y a solubilisation des protéines membranaires et des phospholipides de la membrane
Fig 10-25 • SDS : détergent anionique • Triton X-100 : détergent ionique • Portion hydrophobe du détergent en vert • Portion hydrophile du détergent en bleu
SDS - Page • Solubilisation des protéines par SDS : solubilisation de la protéine avec dénaturation (déroulement) • Parfois réversible • Electrophorèse en Gel de PolyAcrylamide (PAGE) • A révolutionné l'étude des protéines membranaires
Fig 10-26 • Utilisation de détergents légers pour solubiliser, purifier et reconstituer des systèmes de protéines fonctionnels • Très bel exemple
D - Les globules rouges • = Hématies = érythrocytes • Facilement disponibles • Faciles à isoler • Pas de noyau, pas d'organite interne • Une seule membrane = membrane plasmique • Création de fantômes membranaires
Fig 10-27 Globules rouges en microscopie électronique à balayage
Fig 10-28 • Préparations de fantômes de globules rouges • refermés ou non • retournés ou non
Méthodes • Marquage "vectoriel" : marqueur radioactif ou fluorescent soluble (ne se fixe que sur la face exposée) • Enzymes protéolytiques • Anticorps marqués • SDS-PAGE
Résultats • Certaines protéines traversent la bicouche lipidique • La composition des deux feuillets lipidiques est différente
Fig 10-29 • SDS-PAGE des protéines de la membrane du globule rouge humain environ 15 protéines majeures (15000 à 250000 D) • Spectrine + glycophorine + bande 3 > 60 % en masse des protéines membr anaires
a- Spectrine • Protéine du cytosquelette associée à la face cytosolique de la membrane du globule rouge • Principal constituant du cytosquelette qui donne sa forme biconcave au GR • Permet au GR de se déformer pour passer dans les petits capillaires • Mutations dans le gène de la spectrine anémie avec GR sphériques (=sphérocytose héréditaire = Minkowski-Chauffard) • 25 % en masse des protéines associées à la membrane • Longueur = 100 nm (250 000 copies / cellule)
Fig 10-30 A • Molécules de spectrine du GR humain • Hétérodimère qui forme des tétramères • deux chaînes polypeptidiques et anti parallèles enroulées • extrémité phosphorylée pour former le tétramère
Fig 10-30 B Molécules de spectrine du GR humain en microscopie électronique
100 nm Fig 10-31 A • Hétérodimère qui forme des tétramères (200 nm de long) • 4 ou 5 tétramères sont liés dans des complexes de jonction qui contiennentactine(13 monomères)bande 4.1, adducineettropomyosine
Liaison de spectrine à la membrane • Ankyrine • Liaison de la spectrine à la membrane • Se lie à • spectrine • bande 3 (protéine transmembranaire) • Limite la diffusion latérale de bande 3 • Bande 4.1 • Liaison de la spectrine à la membrane • Se lie à • spectrine et ankyrine • bande 3 et glycophorine
Fig 10-31B • Spectrine du cytosquelette de la face cytosolique du GR humain en microscopie électronique (coloration négative)
Autres cellules que le GR • Beaucoup plus compliqué • Cortex du cytosol riche en actine • Cf. chapitre cytosquelette
b - Glycophorine Une des premières protéines membranaires à être séquencée 1 million de molécules par cellule 131 acides aminés Fonction inconnue N'existe que dans le GR Type de la molécule transmembranaire à un passage
Glycophorine • En général homodimère • Extrémité -N à l'extérieur • 100 résidus glucose (90 % des sucres de la membrane du GR) • Un passage : hélice hydrophobe de 23 acides aminés
Fig 10-32 • Transformation d'une chaîne protéique à plusieurs passages en deux chaînes protéiques à plusieurs passages • A et B = même résultat • A : un gène • B : deux gènes
c - Bande 3 • Protéine transmembranaire de 930 acides aminés • 12 passages • Transport du CO2 des tissus vers les poumons • Transporteur d'anions qui permet à HCO3- de traverser la membrane • Se voit en cryofracture
Photo de Branton Daniel Branton (1932 - )
Fig 10-33 • Congélation dans l'azote liquide • Fracture du bloc de glace avec un couteau • Le plan de fracture passe entre les deux feuillets hydrophobes de la membrane • Les plans de fracture sont ombrés au platine • Examen de la réplique au microscope électronique à transmission