Download

1 / 36
431 likes | 1.81k Vues
Electron Transport System: The Chemiosmotic Theory, redox reactions of the electron transport system, NAD/ATP exchange ratio . Bioc 460 Spring 2008 - Lecture 29 (Miesfeld). Passing the baton is analogous to passing along the e- in the ETS.

E N D
Electron Transport System:The Chemiosmotic Theory, redox reactions of the electron transport system, NAD/ATP exchange ratio Bioc 460 Spring 2008 - Lecture 29 (Miesfeld) Passing the baton is analogous to passing along the e- in the ETS Hydrogen cyanide is a deadly gas that blocks e- transport from complex IV to O2 in the ETS Rotenone (rat poison) blocks e- transport through FeS clusters
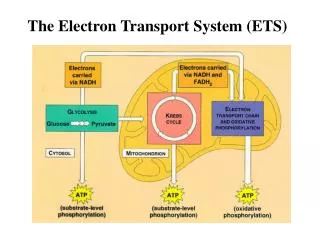
Key Concepts in the Electron Transport System • The Electron Transport System converts redox energy into proton-motive force. In this “pathway” the oxidation of NADH and FADH2 is coupled to the reduction of O2 to form H2O. The proton motive force is used to induce conformational changes in the ATP synthase complex. • The Chemiosmotic Theory states that energy from redox reactions is translated into vectorial energy by coupling electron transfer to membrane bound proton pumps that transverse a proton impermeable membrane and thereby establish an electrochemical proton gradient. • The ATP currency exchange ratios of NADH and FADH2 reflect ATP synthesis in response to H+ movement through the ATP synthase complex. It takes 4 H+ to synthesize 1 ATP, and since NADH oxidation pumps across 10 H+, the exchange ratio is 2.5 ATP/NADH. However, FADH2 oxidation only results in 6 H+ being pumped across the membrane, and therefore the exchange ratio is 1.5 ATP/ FADH2.
The Electron Transport System (ETS) is intimately linked to the process of oxidative phosphorylation, both of which take place within the mitochondrial matrix. Photosynthesis also uses a form of electron transport that is driven by light absorption rather than redox energy. Oxidation of reduced carbon molecules obtained from carbohydrates, lipids, and amino acids feed into the citrate cycle and the ETS.
Peter Mitchell's Chemiosmotic Theory • Oxidation of NADH and FADH2 in the mitochondrial matrix by the electron transport system links redox energy to ATP synthesis. • The chemiosmotic process was first proposed in 1961 by Peter Mitchell, a eccentric British biochemist. Mitchell’s idea was denounced by scientists for 20 years. Mitchell received the 1978 Noble Prize for his breakthrough thinking in energy conversion. • Chemiosmosis involves the outward pumping of H+ from the mitochondrial matrix through three protein complexes in the electron transport system (ETS complexes I, III, IV). • H+flow back down the proton gradient through the membrane-bound ATP synthase complex in response to a chemical (H+ concentration) and electrical (separation of charge) differential.
Overview of Chemiosmotic Theory Electron Transport System FADH2 Ox Phos ATP synthase complex
Basic Components of the Chemiosmotic Theory • Energy from redox reactions or light is translated into vectorial energy • coupling of electron transfer to membrane bound proton pumps that transverse a proton impermeable membrane • thereby establishing an electrochemical proton gradient • A "proton circuit" is established • protons respond to the chemical and electrical gradient across the membrane • flow back across the membrane through the ATP synthase protein complex to catalyze ATP synthesis
Basic Components of the Chemiosmotic Theory • Vectorial H+ pumping results in both: • a chemical gradient across the membrane represented by ΔpH • an electrical gradient due to the separation of charge which can be measured as a membrane potential ΔΨ (Δpsi) • Separation of charge is due to: • build-up of positively-charged protons (H+) on one side of the membrane • accumulation of negative charges (OH-) on the other side of the membrane
Basic Components of the Chemiosmotic Theory • In mitochondria, the contribution of ΔΨ(ΔV) to ΔG is actually greater than that of ΔpH (the ΔpH across the mitochondrial membrane is only 1 pH unit) • In chloroplasts, the ΔpH contribution to ΔG is much more significant with ΔpH close to 3 pH units • Change in free energy (ΔG) for a membrane transport process is the sum of the ion concentration (RT·ln(C2/C1)) and the membrane potential (ZFΔV) • In mitochondria, the ZFΔV term makes a larger contribution than does RT·ln(C2/C1).
The Mitochondrion, the Powerhouse of the Cell A critical feature of the mitochondrion is the extensive surface area of the inner mitochondrial membrane which forms the proton-impermeable barrier required for chemiosmosis. Electron microscopy studies have shown that the inner mitochondrial membrane forms structures called cristae which have been estimated to cover as much as 3,000 m2 per cell (~5 m2 per mitochondrion).
Peter Mitchell - Eccentric Scholar He established the Glynn Research Institute in the early 1960s with a research staff of less than twenty, and remained a private research institution for almost 30 years. Mitchell's uncle was Sir Godfrey Mitchell who owned George Wimpy and Company Limited, the largest construction company in England at the time.
How was Mitchell’s idea proven? Using biochemical approaches: • "inside-out" submitochondrial membrane vesicles that could be shown to pump protons into the interior of the vesicle when oxidizable substrate was made available. • artificial vesicles containing bacterial rhodopsin protein were exposed to light • proton pumping by the bacteriorhodopsin protein resulted in both inward proton pumping • ATP synthesis on the vesicle surface
The Nobel Prize in Chemistry 1978"for his contribution to the understanding of biological energy transfer through the formulation of the chemiosmotic theory“ Peter Mitchell's speech at the Nobel Banquet, December 10, 1978: The philosopher Karl Popper, the economist F. A. Hayek, and the art historian K. H. Gombrich have shown that the creative process in science and art consists of two main activities: an imaginative jumping forward to a new abstraction or simplified representation, followed by a critical looking back to see how nature appears in the light of the new vision. The imaginative leap forward is a hazardous, unreasonable activity. Reason can be used only when looking critically back. Moreover, in the experimental sciences, the scientific fraternity must test a new theory to destruction, if possible. Meanwhile, the originator of a theory may have a very lonely time, especially if his colleagues find his views of nature unfamiliar, and difficult to appreciate. The final outcome cannot be known, either to the originator of a new theory, or to his colleagues and critics, who are bent on falsifying it. Thus, the scientific innovator may feel all the more lonely and uncertain. Last, but not least, I would like to pay a most heartfelt tribute to my helpers and colleagues generally, and especially to those who were formerly my strongest critics, without whose altruistic and generous impulses, I feel sure that I would not be at this banquet today.
PathwayQuestions • What does the electron transport system/oxidative phosphorylation accomplish for the cell? • Generates ATP derived from oxidation of metabolic fuels accounting for 28 out of 32 ATP (88%) obtained from glucose catabolism. • Tissue-specific expression of uncoupling protein-1 (UCP1) in brown adipose tissue of mammals short-circuits the electron transport system and thereby produces heat for thermoregulation. • What is the overall net reaction of NADH oxidation by the coupled electron transport and oxidative phosphorylation pathway? 2 NADH + 2 H+ + 5 ADP + 5 Pi + O2→ 2 NAD+ + 5 ATP +2 H2O
Pathway Questions 3. What are the key enzymes in the electron transport and oxidative phosphorylation pathway? ATP synthase complex – the enzyme responsible for converting proton-motive force (energy available from the electrochemical proton gradient) into net ATP synthesis through a series of proton-driven conformational changes. NADH dehydrogenase – also called complex I or NADH-ubiquinone oxidoreductase. This enzyme catalyzes the first redox reaction in the electron transport system in which NADH oxidation is coupled to FMN reduction and pumps 4 H+ into the inter-membrane space. Ubiquinone-cytochrome c oxidoreductase - also called complex III, translocates 4 H+ across the membrane via the Q cycle and has the important role of facilitating electron transfer from a two electron carrier (QH2), to cytochrome c, a mobile protein carrier that transfers one electron at a time to complex IV. Cytochrome c oxidase - also called complex IV pumps 2 H+ into the inter-membrane space and catalyzes the last redox reaction in the electron transport system in which cytochrome a3 oxidation is coupled to the reduction of molecular oxygen to form water ( O2 + 2 e- + 2 H+→ H2O).
Pathway Questions 4. What are examples of the electron transport system and oxidative phosphorylation? Cyanide binds to the heme group in cytochrome a3 of complex IV and blocks the electron transport system by preventing the reduction of oxygen to form H2O. Hydrogen cyanide gas is the lethal compound produced in prison gas chambers when sodium cyanide crystals are dropped into sulfuric acid.
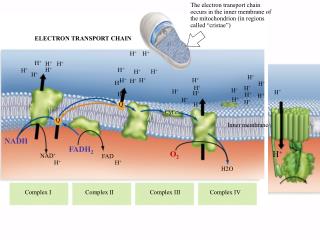
The Electron Transport System Is A Series Of Coupled Redox Reactions The electron transport system consists of five large protein complexes: • Complex I; NADH-ubiquinone oxidoreductase (NADH dehydrogenase • Complex II; succinate dehydrogenase (citrate cycle enzyme • Complex III; Ubiquinone-cytochrome c oxidoreductase • Complex IV; cytochrome c oxidase • F1F0 ATP synthase complex consisting of a "stalk" (F0) and a spherical "head" (F1)
It was possible to order the four electron transport system complexes because of: • Specific redox reaction inhibitors (such as rotenone, antimycin A and cyanide) • Known reduction potentials (Eº') of conjugate redox pairs
The stoichiometry of "proton pumping" is: 4 H+ in complex I 4 H+ in complex III 2 H+ in complex IV (10 H+/NADH and 6 H+/FADH2) FADH2
Metabolic Fuel for Electron Transport • NADH and FADH2 feed into the electron transport system from the citrate cycle and fatty acid oxidation pathways. • Pairs of electrons (2 e-) are donated by NADH and FADH2 to complex I and II, respectively • Pairs of electrons flow through the electron transport system until they are used to reduce oxygen to form water (O2 + 2 e- + 2 H+→ H2O). • The two mobile electron carriers in this series of reactions are coenzyme Q (Q), also called ubiquinone, and cytochrome c which transfer electrons between various complexes.
How is the energy released by redox reactions used to "pump" protons into the inter-membrane space? Answer: we don’t completely know yet, but, it is thought to involve: • a redox loop mechanism • Q cycle in complex III • redox-driven conformational changes : “proton pump” • complexes I and IV
Separation of the H+ and e- on opposite sides of the membrane The Q cycle in complex III uses this mechanism to translocate protons across the membrane
Redox-driven conformational changes in the protein complex "pump" protons across the membrane by altering pKa values of functional groups located on the inner and outer faces of the membrane. Both complexes I and IV have properties that are consistent with such a proton pumping mechanism
Complex I: NADH-ubiquinone oxidoreductase Complex 1 passes 2 e- obtained from the oxidation of NADH to Q using a coupled reaction mechanism that results in the net movement of 4 H+ across the membrane Contains a covalently bound flavin mononucleotide (FMN) that accepts the two electrons from NADH, as well as at least six different iron-sulfur centers (Fe-S) that carry one electron at a time from one end of the complex to the other. The poison rotenone blocks electron transfer within complex I by preventing a redox reaction between two Fe-S centers.
Complex II: Succinate dehydrogenase The citrate cycle enzyme we first encountered in lecture 28. It catalyzes an oxidation reaction that converts succinate to fumarate in a coupled redox reaction involving FAD. The 2 e- extracted from succinate in the citrate cycle is passed through the other protein subunits in the complex to Q as shown below. No protons are translocated across the inner mitochondrial membrane by complex II.
Complex III: Ubiquinone-cytochrome c oxidoreductase The docking site for QH2 (ubiquinol) and consists of 11 protein subunits in each of two monomer subunits. Note the relative position of the electron carriers and the presence of two distinct binding sites for ubiquinone called QP and QN, which play a crucial role in diverting one electron at a time to cytochrome c via the Q cycle. The terms QP and QN refer to the proximity of the sites to the positive (inter-membrane space) and negative (matrix) sides of the membrane.
The Q Cycle(No, it isn’t an invention for James Bond) James Bond Q …the humor of David Gang
Functions as both a mobile electron carrier and a "transformer" that converts the 2 e- transport system used by complexes I and II, into a 1 e- transport system required by cytochrome C. The Q cycle requires that 2 QH2 molecules get oxidized by complex III, with one of QH2 molecule being re-formed by reduction to give a net oxidation of 1 QH2 molecule. The Q Cycle
Four Steps of the Q Cycle Converts a 2 e- carrier (QH2) into a 1 e- carrier (cytochrome c). This requires that 2 QH2 molecules be oxidized, with 1 QH2 being reformed.
2 H+P Note that the Q cycle reactions require that 2H+ from the matrix be used to regenerate QH2, even though 4H+ are translocated. However, this apparent imbalance of 2H+is corrected by the redox reactions of complex IV where2H+ are required to reduce oxygen to waterand 2H+ are pumped across the membrane. Therefore, the net translocation of protons across the membrane in the combined redox reactions of complexes III and IV becomes 6 H+N→6 H+P. 2 H+P 2 H+N
To see how the Q cycle accomplishes the 2 e-→ 1 e- + 1 e- conversion process, write out two separate QH2 oxidation reactions and then sum them to get the net reaction for complex III: QH2 + Cyt c (oxidized) → Q•- + 2 H+P + Cyt c (reduced) QH2 + Q•- + 2 H+N + Cyt c (oxidized) → Q + QH2 + 2 H+P + Cyt c (reduced) QH2 + 2 H+N + 2 Cyt c (oxidized) → Q + 4 H+P + 2 Cyt c (reduced) Why are 2 QH2 required to transfer 2 e- to 2 cytochrome c molecules? What happens to the “extra” QH2 molecule in step 4 of the Q cycle?
Cytochrome C Cytochrome c (Cyt c) is a small protein of ~13 kDa that associates with the cytosolic side of the inner mitochondrial membrane and is responsible for transporting one electron at a time from complex III to complex IV using an iron-containing heme prosthetic group. Oxidized Cyt c contains ferric iron (Fe3+) in the heme group, reduced Cyt c contains ferrous iron (Fe2+). A version of the Cyt c molecular structure is used in the Bioc460 website header. mystery protein revealed
Complex IV: Cytochrome c oxidase 2 H+P Complex IV accepts electrons one at a time from Cyt c and donates them to oxygen to form water. In the process, two net H+ are pumped across the membrane using a conformational-type mechanism similar to complex I. Cyt c docks on the P side of the membrane to complex IV near CuA which accepts the electron leading to oxidation of the heme group in Cyt c (Fe2+ --> Fe3+). Cyanide blocks electron transfer in complex IV. 4 H+N
ATP Currency Exchange Ratios of NADH and FADH2 Experimental measurements demonstrate 3 H+ are required to synthesize 1 ATP when they flow back down the electrochemical proton gradient through the ATP synthase complex, and 1 H+ is needed to transport each negatively-charged Pi molecule into the matrix.
ATP Currency Exchange Ratios of NADH and FADH2 Taking into account the requirement of 3 H+/ATP synthesized, and the use of 1 H+ to translocate ADP, the total is 4 H+/ATP. We can now see where the ATP currency exchange ratios of ~2.5 ATP/NADH and ~1.5 ATP/FADH2 come from: Oxidation of NADH starting at complex I yields: 10 H+/4 H+ = 2.5 ATP Oxidation of FADH2 starting at complex II yields: 6 H+/4 H+ = 1.5 ATP for FADH2