Transcription
Transcription. Why regulation of gene expression is important? Cellular function is largely dictated by the set of macromolecules inside the cell. Different macromolecules accumulate to different levels under different growth conditions and in different cell types.


Transcription
E N D
Presentation Transcript
Why regulation of gene expression is important? • Cellular function is largely dictated by the set of macromolecules inside the cell. • Different macromolecules accumulate to different levels under different growth conditions and in different cell types. • Diseases can be caused by aberrant control of gene expression: too much or too little of a protein; wrong time and wrong place for a protein.
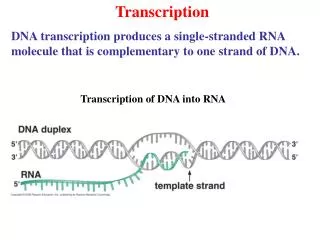
Transcription and translation in eukaryotic cells are separated in space and time.Extensive processing of primary RNA transcripts in eukaryotic cells.
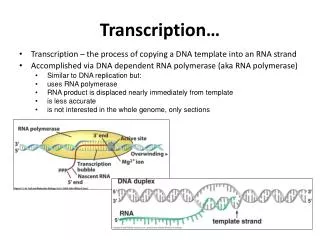
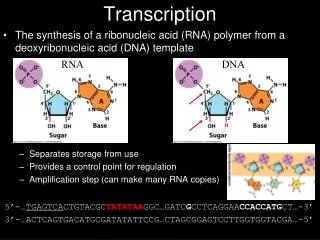
Transcription of DNA into RNA by RNA polymerase---an overview1. Requires DNA template, four ribonucleotide 5’ triphosphates, Mg+2. 2. De novo synthesis: does not require a primer.Low fidelity compared to DNA polymerase: errors 1/104-105 (105 higher than DNA polymerase).3. Activity highly regulated in vivo: at initiation, elongation and termination.4. The nucleotide at the 5’ end of an RNA strand retains all three of its phosphate groups; all subsequent nucleotides release pyrophosphate (PPi) when added to the chain and retain only their a phosphate (red). 5. The released PPi is subsequently hydrolyzed by pyrophosphatase to Pi, driving the equilibrium of the overall reaction toward chain elongation. 6. Growth of the transcript always occurs in the 5’-to-3’ direction. Template (non -coding) strand Non-template (coding) strand
Transcription in Prokaryotes • RNA polymerase from E. coli: A multisubunit structure of the forma2bb’salpha, beta, omega, sigma) • The holoenzyme loses the s subunit to give the core enzyme. • The function of the s unit is to recognize the promoter locus.
Sigma Units in E. coli. • s70 (70 kD) promotes transcription of most genes. • s32promotes transcription of heat shock genes. • promotes transcription of the flagellin gene. • 54 is synthesized to assimilate nitrogen during nitrogen starvation.
E. coli RNA polymerase holoenzyme bound to DNA SubunitStoichiometryRole in holoenzyme a 2 Binds regulatory sequences/proteins Forms phosphodiester bonds/binds ribo- nucleoside triphosphate substrates ’ Promoter recognition RNAP assembly A single RNA polymerase makes multiple types of RNAs (rRNA, tRNA and mRNA) in prokaryotes. Known as Processive.
Typical E.coli promoters recognized by an RNA polymerase holoenzyme containing s70 Strong promoters: frequent initiation of transcription-every 2 seconds Weak promoters: may contain substitutions, transcribed every 10 min.
Biochemical studies of bacterial RNA polymerase DNA binding assay- DNase I footprinting to look for polymerase-bound promoters Role of individual subunits- dissociation of holoenzyme by column chromatography
DNase I footprinting: a common technique for identifying protein-binding sites in DNA.1. A DNA fragment is labeled at one end with 32P (red dot).2. Portions of the sample then are digested with DNase I in the presence and absence of a protein that binds to a specific sequence in the fragment. 3. A low concentration of DNase I is used so that on average each DNA molecule is cleaved just once (vertical arrows). 4. The two samples of DNA then are separated from protein, denatured to separate the strands, and electrophoresed. The resulting gel is analyzed by autoradiography, which detects only labeled strands and reveals fragments extending from the labeled end to the site of cleavage by DNase I.
[NaCl] [protein] ’ ’ Dissociation of RNAP and purification of sby ion-exchange chromatography Absorbance at 280 nm Carboxymethyl- (-CO2-2) or phospho- (-PO3-2) cellulose Fraction number
’ The dissociable sigma subunit gives promoter specificity to prokaryotic RNA polymerase (RNAP) + ’ Core enzyme Holoenzyme Specific promoter binding; weak non-specific DNA binding (Kd ~10-7 M); finds promoter 10,000 times faster. No specific promoter binding; tight non-specific DNA binding (Kd ~5 x 10-12 M)
Initiation: Consensus Sequences • In prokaryotes: often 25 and 10 base pairs upstream of the start of transcription -35 box and –10 (Pribnow) box TTGACA TATAAT +1 will be the start site • In eukaryotes: TATA box lies 25 base pairs upstream
Initiation: Promoter Locus • Promoter locus: A DNA segment that signals the start of RNA synthesis. • Is upstream (toward the 3’ end) of the DNA segment where the gene coding for the RNA actually begins. • Prokaryote promoter regions contain many sequences in common called consensus sequences. These are rich in A-T base pairs
’ ’ ’ Holoenzyme Transcription initiation by prokaryotic RNA polymerase “sliding and scanning” Promoter -35 -10 Closed complex Sigma separates from the core once a few phosphodiester bonds are formed rNTPs PPi Core enzyme Open complex; initiation 5’pppA/G mRNA
Interactions of various sigma factors of E. coli with the same core polymerase to form holoenzymes with different promoter-binding specificity SigmaFactor Promoters RecognizedPromoter Consensus-35 Region -10 Regions70 Most genes TTGACAT TATAATs32 Genes induced by heat shock TCTCNCCCTTGAA CCCCATNTAs28 Genes for motility and chemotaxis CTAAA CCGATATs38 Genes for stationary phase and stress response ? ? -24 Region -12 Regions54 Genes for nitrogen metabolism & other functions CTGGNA TTGCA Heat-shock response: High temperature induces the production of s32, which binds to the core polymerase to form a unique holoenzyme for recognition of the promoters of heat-shock induced genes.
Elongation • The s unit leaves after about 10 nucleotides have added to the RNA. • Core polymerase binds accessory proteins and RNA synthesis continues in the 5’→3’ direction. • The transcription “bubble” (DNA-RNA hybrid) leaves positive supercoils ahead of and negative supercoils behind on the DNA as it moves “downstream”.
Transcriptional elongation: Movement of transcription bubble (17-bp, 1.6 turns of B-DNA duplex) Speed of movement: 50-90-nt/sec Supercoiling of DNA during transcription causes a requirement for topoisomerases
Termination (Prokaryotes) Two forms of prokaryotic termination: • Termination sequences contain palindromes (a palindrome is a word ,phrase or sentence that is spelled identically read either forward or backwad;two examples are ROTATOR and NURSES RUN. The term is applied to regions of DNA with inverted repeats of base sequence having twofold symmetry over two strands of DNA).The RNA transcript forms a hairpin turn which disrupts the DNA-RNA hybrid. (Rho-independent) • In r-dependent terminmation, the r protein binds to RNA and catlayzes dissociation of the poylmerase from the DNA-RNA hybrid. (Rho-dependent)
Palindromic DNA 5’--- T T A G C A C G T G C T A A--- 3’ 3’--- A A T C G T G C A C G A T T--- 5’ 5’------TGCGATACTGATCGCA-------3’ 3’------ACGCTATGACTAGCGT-------5’ RNA : 5’------UGCGAUACUGAUCGCA------3’
Rho-independent prokaryotic transcription termination The core polymerase pauses after synthesizing a hairpin. If the hairpin is really a terminator, RNA will dissociate from the DNA strand as the A-U pairing is unstable. Once the RNA is gone, DNA duplex reforms and the core is driven off, as it has low affinity for dsDNA.
Rho-dependent transcription termination Rho-binding Site (non-contiguous structural features in RNA): Stop signals not recognized by RNAP alone. Rho: forms RNA-dependent hexameric ATPase, translocates along RNA 5’-to-3’, pulling RNA away when it reaches the transcription bubble. termination Platt, Ann. Rev. Biochem.55: 339 (1986)
Termination (Prokaryotes)-2 • mRNA is used immediately. • Mature rRNA and tRNA are processed from larger transcripts. tRNA bases are frequently modified after transcription. Unusual bases are popular in tRNAS. Except the four common bases ( A, U, G, C ), the other bases are called unusual bases, such as pseudouridine , dihydrouridine and so on
Modification of RNAs • 1. Leader and trailer sequences are removed. (Trimming) • 2. Terminal sequences can be added. • 3. Base modification can occur.
Modification of RNAs: tRNA • Base modification occurs both before and after trimming. Methylation and substituting S for O are common. • Often several tRNAs are synthesized in one long chain which is split into fragments after synthesis. • RNase P makes 5’ ends of all E. coli tRNAs. The enzyme consists of both protein and RNA with the RNA being catalytically active.
Modification of RNAs: tRNA-2 • All tRNAs have a CCA sequence at the 3’ end. This end is where the amino acid to be transferred to the ribosome is attached. • tRNAs have a three-dimensional L shape as shown on slide 28 for yeast alanyl tRNA.
Attachment to mRNA here AA attaches here Stem region tRNA: cloverleaf; “L” model
Modification of RNAs: rRNA-1 • rRNA processing consists mainly of methylation and trimming to proper size. • There are three rRNAs in a prokaryotic ribosome and four in eukaryotic ribosomes. The next slide summarizes rRNA processing in E. coli. Prokaryote: 23s ,16s , and 5s rRNAs Eukaryote: 28s, 18s, 5.8s , and 5s rRNAS s: Svedberg unit , represents the molecular size of a RNA
Modification of RNAs: rRNA-2 Individual RNases are identified by letters/numbers, e. g. M5, X and III. Prokaryotic rRNA
Eukaryotic rRNA 28s, 18s , and 5.8s rRNAs are transcribed by RNA polymerase I , exist on the same primary rRNA transcript, and processed by ribonucleases to produce three different rRNAs. 5s rRNA is transcribed by RNA polymerase III and modified separately.
Modification of RNAs: mRNA • Prokaryotic mRNA is used as synthesized. • Eukaryotic mRNA is processed extensively: • 1. Capping occurs at the 5’ end. • 2. Polyadenylation occurs at the 3’ end. • 3. Splicing of code sequences (called exons) occurs and noncoding sequences (introns) are removed.
Transcription (Eukaryotes) 1. The three RNA polymerases differ in the type of RNA synthesized, subunit structure, and relative amounts. a.) RNA Pol I transcribes large rRNA. b.) RNA Pol II transcribes precursors to mRNA (hn RNA,heterogenous nuclear RNA) and most snRNAs. c.) RNA Pol III transcribes precursors of tRNAs and 5S rRNA.
Inhibition of α-amanitin • RNA polymerase II is inhibited by α-amanitin---a toxic polypeptide produced by poisonous mushroom Amanita phalloides (sometimes called“death cap” or “destroying angel”---it is said to taste delicious and very beautiful)
Upstream control element Core promoter element TATA-box GC-box CCAAT-box Repressor Initiator Core promoter 5S RNA tRNA U6 RNA Box A Box C Box A Box B TTATATAT-box Proximal sequence element octamer Three types of Eukaryotic promoters: • RNA polymerase (pol) I synthesizesthe large rRNA • RNA polymerase (pol) II synthesizes mRNA • Inducible • Repressive • tissue-specific gene expressions via binding to their target sequences • RNA polymerase (pol) III synthesizestRNA and 5S rRNA. Modified from: Tamura T.-A. et al., 1996
Transcription ( Eukaryotes )-2 2. Promoters are larger and more complex. a.) Often the TATA box 25-30 bp upstream promotes transcription by Pol II. b.) Various complexes form, the DNA strands unwind at the TATA region, and the process begins. c.) a lot of transcription factors (TF) are involved.
Transcription (Eukaryotes)-3 3. Processing is more complex for eukaryotic mRNA. hnRNA is associated with nuclear proteins in ribonuclearprotein particles (hnRNP). a.) Cap structure protects 5’ end from exo-nucleases. (Next slide) b.) The 3’ end is chopped and 100-250 adenylate residues are added to protect the end and promote export of mRNAs.
First step in RNA processing: CappingFormation of the 7-methylguanine 5’- 5’ cap structure. The 5’ cap structure is essential for efficient pre-mRNA splicing, export, stability and translation initiation.Three separate enzyme activities are required for cap formation.PhosphataseGuanyl transferaseMethyl transferaseThe cap protects the RNA from 5’-exonucleolytic cleavage *Note the unusual 5’ – 5’ linkage *Note the 7-methylguanine
Exons, Introns, and Splicing • Eukaryotic genes often contain base sequences that do not appear in the mature mRNA expressed by the gene. These sequences are called introns. They are cut from the RNA nucleotide chain and the exons (the bases that are expressed) are spliced together. • The number of introns in a gene can vary from none to many (>50).
Exons, Introns, and Splicing-2 • Introns are removed in the nucleus by small nuclear ribonuclearproteins, snRNPs. (pronounced ‘snurps’) • snRNPs function by cutting the mRNA, forming a lariat (loop) of the introns, and joining the two pieces of exon as the loop is cleaved. See the next slide. • Some RNAs catalyze their own self-splicing. • Lupus results from antibodies to a snRNP.
Exons, Introns, and Splicing-3 • Splicing begins with nucleophillic attack of the 2’-OH on A on the 5’ splice site. • A 2’-5’ phospho-diester bond forms the lariat. • The 3’-OH attacks the phosphate next to the lariat.
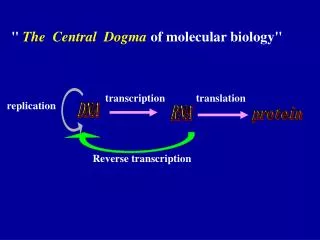
Reverse transcription • Retrotranscription( Reverse transcription) DNA is synthesized by Using RNA strand as the template ,the reaction is catalyzed by reverse transcriptase. extension of central dogma