Download

1 / 29
320 likes | 595 Vues
Protein Turnover and Amino Acid Catabolism. Pepsin nonspecific maximally active at low pH of the stomach. Proteolytic enzymes of the pancreas in the intestinal lumen display a wide array of specificity. Aminopeptidases digest proteins from the amino-terminal end.

E N D
Pepsin nonspecific maximally active at low pH of the stomach. • Proteolytic enzymes of the pancreas in the intestinal lumen display a wide array of specificity. Aminopeptidases digest proteins from the amino-terminal end. The Digestion and Absorption of Dietary Proteins
Cellular Proteins Are Degraded at Different Rates • Some proteins are very stable, while others are short lived. • Altering the amounts of proteins that are important in metabolic regulation can rapidly change metabolic patterns. • Cells have mechanisms for detecting and removing damaged proteins. • A significant proportion of newly synthesized protein molecules are defective because of errors in translation. • Other proteins may undergo oxidative damage or be altered in other ways with the passage of time.
Ubiquitin Tags Proteins for Destruction • How can a cell distinguish proteins that are meant for degradation? • Ubiquitin, a small (8.5-kd) protein present in all eukaryotic cells, is the tag that marks proteins for destruction.
The c-terminal glycine residue of ubiquitin (Ub) becomes covalently attached to the e-amino groups of several lysine residues on a protein destined to be degraded. • The energy for the formation of these isopeptidebonds (iso because e- rather than a-amino groups are targeted) comes from ATP hydrolysis.
Three enzymes participate in the attachment of ubiquitin to each protein: • ubiquitin-activating enzyme, or E1 • ubiquitin-conjugating enzyme, or E2 • ubiquitin-protein ligase, or E3.
Chains of ubiquitin can be generated by the linkage of the e-amino group of lysine residue 48 of one ubiquitin molecule to the terminal carboxylate of another. • Chains of four or more ubiquitin molecules are particularly effective in signaling degradation
What determines whether a protein becomes ubiquitinated? • The half-life of a cytosolic protein is determined to a large extent by its amino-terminal residue “the N-terminal rule”. • A yeast protein with methionine at its N terminus typically has a half-life of more than 20 hours, whereas one with arginine at this position has a half-life of about 2 minutes. • A highly destabilizing N-terminal residue such as arginine or leucine favors rapid ubiquitination, whereas a stabilizing residue such as methionine or proline does not. • E3 enzymes are the readers of N-terminal residues. • Cyclin destruction boxes are amino acid sequences that mark cell-cycle proteins for destruction. • Proteins rich in proline, glutamic acid, serine, and threonine (PEST sequences).
The Proteasome Digests the Ubiquitin-Tagged Proteins • A large protease complex called the proteasome or the 26S proteasome digests the ubiquitinated proteins. • This ATP-driven multisubunit protease spares ubiquitin, which is then recycled. • The 26S proteasome is a complex of two components: • 20S proteasome, which contains the catalytic activity1 • 19S regulatory subunit.
The OH groups of these aas are converted into nucleophiles with the assistance of their own amino groups. • These nucleophilic groups then attack the carbonyl groups of peptide bonds and form acyl-enzyme intermediates. • ATP hydrolysis may assist the 19S complex to unfold the substrate and induce conformational changes in the 20S proteasome so that the substrate can be passed into the center of the complex
P P E3 P P Ub NF-kB NF-kB I-kB P P Ub Ub P Ub P Ub proteosome Protein Degradation Can Be Used to Regulate Biological Function Example: E3 Inflammation initiates the expression of a number of the genes that take part in this response
Digested proteins Amino Acids Degradation in the liver NH4+ A-ketoacids The amino group must be removed, as there are no nitrogenous compounds in energy-transduction pathways enter the metabolic mainstream as precursors to glucose or citric acid cycle intermediates
The fate of the a-amino group • The a-amino group of many aas is transferred to a-ketoglutarate to form glutamate. • Glutamate is then oxidatively deaminated to yield ammonium ion (NH4+).
Aminotransferases (transaminases) catalyze the transfer of an a-amino group from an a-amino acid to an a-keto acid.
Example: • Aspartate aminotransferase: • Alanine aminotransferase: • These transamination reactions are reversible and can thus be used to synthesize amino acids from a-ketoacids,
The nitrogen atom that is transferred to a-ketoglutarate in the transamination reaction is converted into free ammonium ion by oxidative deamination. • This reaction is catalyzed by glutamate dehydrogenase. • This enzyme is unusual in being able to utilize eitherNAD+ or NADP+ at least in some species. • The reaction proceeds by dehydrogenation of the C-N bond, followed by hydrolysis of the resulting Schiff base.
Glutamate dehydrogenase and other enzymes required for the production of urea are located in mitochondria. • This compartmentalization sequesters free ammonia, which is toxic. • In most terrestrial vertebrates, NH4+ is converted into urea, which is excreted.
Pyridoxal Phosphate Forms Schiff-Base Intermediates in Aminotransferases • All aminotransferases contain the prosthetic group pyridoxal phosphate (PLP), which is derived from pyridoxine (vitamin B6).
Pyridoxal phosphate derivatives can form stable tautomeric forms The most important functional group allows PLP to form covalent Schiff-base intermediates with amino acid substrates a pyridine ring that is slightly basic A phenolic hydroxyl group that is slightly acidic
The aldehyde group of PLP usually forms a Schiff-base linkage with the e-amino group of a specific lysine residue of the enzyme. • The a-amino group of the amino acid substrate displaces the e-amino group of the active-site lysine residue.
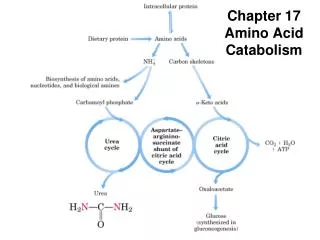
The Urea Cycle • Some of the NH4+ formed in the breakdown of amino acids is consumed in the biosynthesis of nitrogen compounds. • In most terrestrial vertebrates, the excess NH4+is converted intourea and then excreted. • The urea: • One nitrogen atom is transferred from aspartate. • The other nitrogen atom is derived directly from free NH4+ . • The carbon atom comes from HCO3-.
The Urea Cycle Reactions • Formation of Carbamoyl Phosphate: catalyzed by carbamoyl phosphate synthetase. • The consumption of two molecules ofATP makes the synthesis essentially irreversible. • The carbamoyl group of carbamoyl phosphate has a high transfer potential because of its anhydride bond.
Carbamoyl is transferred to ornithine to form citrulline. • The reaction is catalyzed by ornithine transcarbamoylase. • Ornithine and citrulline are amino acids, but they are not used as building blocks of proteins.
Citrulline is transported to the cytoplasm where it condenses with aspartate to form argininosuccinate • The reaction is catalyzed by argininosuccinate synthetase. • The reaction is driven by the cleavage ofATP into AMP and PPi, and by the subsequent hydrolysis ofPPi.
Argininosuccinase cleaves argininosuccinate into arginine and fumarate. • Thus, the carbon skeleton of aspartate is preserved in the form of fumarate.
Arginine is hydrolyzed to generate urea and ornithine in a reaction catalyzed by arginase. • Ornithine is then transported back into the mitochondrion to begin another cycle.
Mitochondrial reactions: • The formation of NH4+ by glutamate dehydrogenase. • Its incorporation into carbamoyl phosphate • Synthesis of citrulline • Cytosolic reactions: • The next three reactions of the urea cycle, which lead to the formation of urea, take place in the cytosol.