Download

1 / 37
370 likes | 382 Vues
Protein and amino acid degradation. Chemistry 256. Proteins don’t last forever; in fact, some are quite short-lived. Thus there must be a mechanism for the cell to recycle the “ broken ” protein ’ s amino acids and not have to synthesize new amino acids, which is energetically more expensive.

E N D
Protein and amino acid degradation Chemistry 256
Proteins don’t last forever; in fact, some are quite short-lived • Thus there must be a mechanism for the cell to recycle the “broken” protein’s amino acids and not have to synthesize new amino acids, which is energetically more expensive. • Shortest-lived enzymes tend to be the ones that catalyze reactions at metabolic control points.
Ways to mark proteins for degradation • In the cell, there are a couple of ways to target damaged or unwanted proteins: 1. Non-selectively, the protein interacts with a lysosome, with its numerous proteases. The process can be selective when energy levels are low and “KFERQ” proteins are targeted. 2. Selectively, the protein is ubiquinated – that is, it is covalently bonded to ubiquitin. This allows recognition by a proteasome, which degrades the protein.
Lysosomes are vesicular structures Lysosomes maintain an internal pH of 5 and contain about 50 hydrolytic enzymes that work optimally at that pH (are inactive at cytosolic pH). The “food vacuole” shown in the diagram probably contains proteins captured by the cell by endocytosis; generally, the process of which proteins are degraded is nonselective, except in the case of starvation, in which case proteins with the sequence KFERQ (lys-phe-glu-arg-gln) or similar are removed selectively for energy purposes from certain cells.
Ubiquitin marks proteins for degradation without the use of a lysosome • Ubiquitin is a 76-residue protein, highly conserved over many species. • Uses ATP and ubiquitin-activating and ubiquitin-conjugating enzymes, as well as a ligase, to covalently link ubiquitin to the protein at a lysine side chain. • Need at least four ubiquitins for protein to be efficiently degraded by the 26S proteasome = polyubiquitin
“N-end rule” • “destabilizing” N-terminus residues = asp, arg, leu, lys, phe – proteins with these N-terminus residues have a half-life of 2 to 3 minutes. • “stabilizing” N-terminus residues = ala, gly, met, ser, thr, val – proteins with these N-terminus residues have a half-life of greater than 10 hours in prokaryotes, greater than 20 hours in eukaryotes (these proteins are basically never ubiquitinized).
Proteasome = degrades ubiquinated proteins • Multiprotein complex – the 20S proteolytic part and the two 19S “end caps” which recognize the ubiquinated proteins. The caps are also ATPases, which provides the free energy to break up the proteins.
The 20S proteasome is not found in eubac-teria; self-compartmentalized proteases are • E. coli’s Clp protease has D7 symmetry Hollow space in the center is the active site; design is to keep protease from indiscriminate destruction (no ubiquitin signal) Side view Top view
Transamination = transfer of amino acid amine to an α-keto acid • Once free amino acids are made (by proteasomes, pepsin, trypsin, whatever) and transported in the bloodstream, they are transaminated in the cell to yield…a different α-keto acid and a different amino acid. • So why bother? The goal is to get to glutamate, which can be deaminated, and the resulting nitrogen excreted safely.
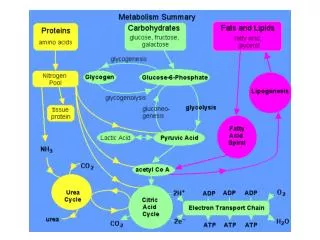
The recovery of carbon and nitrogen from amino acids • The amine enters the urea cycle, and the nitrogen eventually excreted. • The rest of the acid skeleton is recycled in a number of ways.
Schiff base (aka “imine”) Imines are created when a carbon of a ketone or aldehyde is attacked by a (nucleophilic) amine; as shown above, the inter-mediate is an amine with an alcohol on the α-carbon, which will eventually rearrange into a C=N bond. This final product is the imine, or Schiff base. Note the similarity of the first step to hemiketal or hemiacetal formation.
The lysine of the transaminase enzyme forms a Schiff base with the coenzyme pyridoxal-5’-phosphate (PLP) • PLP is derived from vitamin B6. • Snell, Braunstein and Meztler (1954) showed the mechanism of transaminase is “Ping-Pong”.
Mechanism of transamination Note enzyme is displaced by the amino acid but Schiff base remains. Thus the “ammonia” is carried safely.
Selectivity of the transimination In the previous diagram, note that bonds a, b or c can be cleaved (in this diagram, the C-X, C-Y and C-Z bonds to the α-carbon). Which one gets cleaved is a function of which one lies parallel to the π-bond of the imine that is about to form.
Mechanism of transamination • Enzyme hangs around to act as a base.
Mechanism of transamination • Release of the new α-keto acid. • Then what? Do the whole thing again, but backwards to yield either aspartate or glutamate. • Lysine is the only acid that is not transaminated.
Presence of transaminases in the bloodstream is a bad sign • Comes from damaged liver or muscle tissue. • Serum glutamate-oxaloacetate transaminase (SGOT) or serum glutamate-pyruvate transaminase (SGPT) are measured.
Glutamate dehydrogenase deaminates glutamate • Activated by ADP and NAD+ • Inhibited by GTP and NADH • The ammonium ion liberated is eventually excreted through the urea cycle
How to remove carbons: Tetrahydrofolate (THF) is a one-carbon carrier in amino acid degradation
The group that THF carries determines the oxidation state of THF The oxidation state will determine the potential of the reduction needed to transfer the carbon group.
oxidation! oxidation!
So an antibiotic strategy is to introduce a molecule that looks like THF but will not carry a carbon group!
The urea cycle • Hans Krebs & Kurt Henseleit, “Studies on urea formation in the animal organism” Hoppe-Seylers Z. Physiol. Chem. (1932). • First metabolic cycle elucidated. • Purpose: excrete excess nitrogen by converting ammonia into less-toxic urea. • Cost: 3 ATP
Pre-urea cycle step occurs in mitochondrion Multi-step synthesis begins with ATP reacting with bicarbonate ion to form carboxyphosphate, which reacts with ammonia to yield carbamate. Urea cycle occurs mostly in liver, and some in kidneys.
Carbamoyl phosphate synthetase catalyzes step 1 • Enzyme complex is like an assembly line, with a 96 Å long tunnel that allows the substrate to move through as it is processed. • Protects intermediates with short half-lives (“channeling”). • Also concentrates ammonia to make carbamate.
Several different CPS enzymes • CPS I is found in the mitochondrion and catalyzes the precursor step to the urea cycle • CPS II is found in the cytosol and catalyzes pyrimidine metabolism • CPS III is found in fish, and is considered to be an evolutionary-link between glutamine-dependent and ammonia-dependent CPS enzymes
The urea cycle occurs in both mitochonrion and cytosol • Citrulline and ornithine are the components transported in and out of mitochondrion. • Cytochrome c, NADH, ATP are necessary reactants (Philip Cohen, 1946) • Citrulline/aspartate step (Sarah Ratner, 1949).
Urea cycle regulated by substrate concentration • The rate determining step of the urea cycle is the first one – the synthesis of carbamoyl phosphate – so the enzyme carbamoyl phosphate synthase is activated by the presence of N-acetylglutamate, which is produced when there is an excess of glutamate due to protein breakdown.
Amino acid degradation • 10–15% of the amino acids are broken down to CO2 and H2O and the free energy harvested. • The rest enter the TCA cycle in one of two ways: • Glucogenic amino acids are degraded to pyruvate or other TCA intermediate that will become glucose. • Ketogenic amino acids are degraded to acetyl-CoA or acetoacetate and thus can be made into fatty acids or ketone bodies.
Some amino acids can never recovered as glucose • Leucine and lysine degrade to only acetyl-CoA and/or acetoacetate, so can’t become glucose. • Animals have no way of converting either acetyl-CoA or acetoacetate back into a glucose precursor.
Methionine leads to homocysteine • Methionine’s breakdown leads through S-adenosylmthionine to homocysteine, which is a cysteine with an extra methylene on the side chain. • Excess levels of homocysteine have been correlated to cardiovascular disease and the probable mechanism is oxidative damage to arterial walls.
THF is a one-carbon carrier • Note that R in the diagram below is the p-aminobenzoic acid and the glutamate moieties. • The one-carbon units can be methyl, methylene, formyl, formimino, methenyl, and are covalently bonded to N-5 and/or N-10. • Critical in formation of neural tubes, so folic acid is a critical nutrient for moms.